
Why is creation research a growing and developing field of interest? Due to the vast differences between naturalistic, evolutionary and biblical worldviews, the need for such research is significant, as Todd Wood explains in his 2013 review of creation research. Creationists use a biblical worldview to understand origins and the world around us in the context of the Bible.
The following is a summary of “A Review of the Last Decade of Creation Biology Research on Natural History, 2003-2012” by Dr. Todd Wood, and of the surrounding discussion and research pertaining to it. The views expressed do not necessarily reflect those of New Creation.
Genesis gives an account from God about how He made all things in six days and what happened to the world following that week. Creation research seeks to explain the world around us by the catastrophes the Bible teaches about (e.g. the Flood and Babel). Scientists can do this through studies in:
- Biosystematics (to understand created kinds)
- Speciation (understanding the diversity of those created kinds)
- Biogeography (learning how animals have moved and filled the earth)
- Natural evil and pathology (what are the origins of certain features in a fallen world?)
- Biological similarity and design
Biosystematics -Meet Baraminology
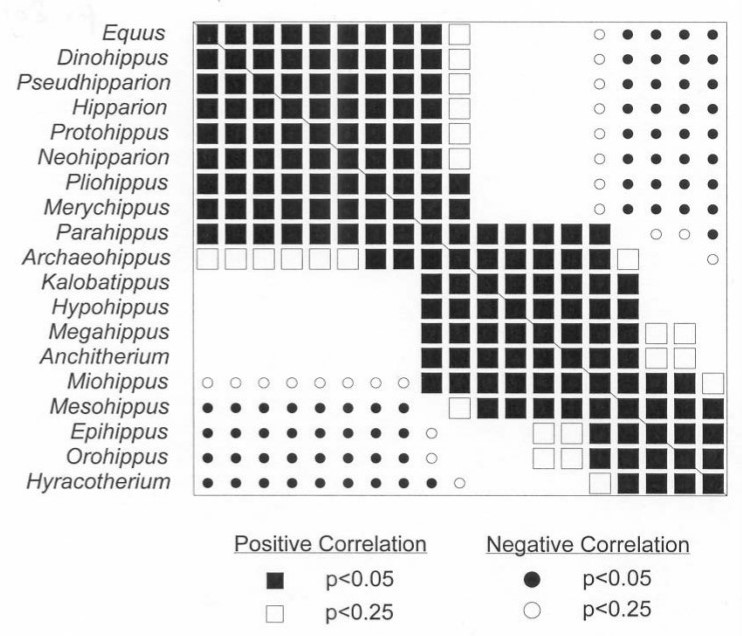
Baraminology, the study of created kinds, asks questions about what organisms God originally created and which organisms today are related within those kinds. Pioneered by Frank Marsh in 19471, baraminology looks at the similarities and discontinuities between organisms to assess their relationships. The data is carefully analyzed using statistical analyses and corrective techniques to keep errors low (e.g. bootstrapping and multidimensional scaling2). Important groups have been studied with this method, such as Hominidae (the family to which humans belong) and Equidae (the family of horses).3, 4 While evolution explains relationships within these families as evidence of a common ancestral descent, baraminology suggests that these may represent created kinds.
 - Own work, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=11633334")
Further Reading on Baraminology:
- Netted Hierarchy of Biological Similarity
- Bombardier Beetles & Baraminology
- Baraminology
- Cenozoic Mammals After Their Kinds
- How Many Kinds of Mammals Were on the Ark?
Speciation
In addition to understanding which organisms are related by God’s original design, Wood explores in his review how studies of speciation can explain how animals and plants have diversified over time. There are many possible ways for species to form and adapt. Some examples include allelic diversity, karyotypic changes, and mutations.
- Allelic diversity (suggested by Byron Nelson in 19275 and Tinkle in 19676) is one possibility in which God created organisms to originally have heterozygosity in their genes. This would allow for great diversity as animals reproduced and hybridized. However, this alone would not explain all the species we see today.
- Changes in species karyotypes (the arrangement and number of chromosomes a species has in its DNA) can play a large role in diversity within created kinds, as found by Lightner.7 The way DNA is packaged and arranged profoundly affects which genes we express. Sometimes parts of our DNA can move around (these parts are called transposons) and can be expressed differently. With these variations, diversity can increase and species can develop from the original organisms God made.
- Mutations are another way in which God may have allowed for great diversity to take place within created kinds. Even a change in one nucleotide of a gene can cause an expression to vary (these Single Nucleotide Polymorphisms, or SNPs, are often thought to be predictors for certain traits like hair or eye color).
Further Reading on Speciation:
- What Is Disparity, and Where Did It Come From?
- What is the eKINDS Project?
- Mediated Design Versus Micro-Evolution
Biogeography
Biogeography is a field which asks questions about where animals and plants came from and how they arrived at their current locations. Creationists are especially interested in this field due to the challenges regarding Flood models. For example, since the continents moved great distances during the Flood due to plate tectonics, how did the animals disperse from the Ark and reach places like North America? Some suggestions involve floating rafts of vegetation and debris which can carry animals overseas.8 Storms and strong winds may also carry plants and animals, and there are even modern examples of this happening.9
Natural Evil and Pathology
Natural evil is an important and complex area of study. God describes the pre-Fall world as “good” in Genesis 1-2, and most biblical creationists believe no death of animals or humans occurred before the Fall. This understanding is due to the Hebrew words for “life” and “death” and how they apply to humans and animals apart from plants.10, 11
Apart from natural death, creationists ask questions regarding predators, parasites, toxins, and pathogens. Where did special adaptations for predation come from if all animals were originally all herbivorous in diet (Genesis 1:29)? Perhaps God specifically designed these features for an original purpose which has since changed after the Fall.12, 13 The role of bacteria and various pathogens may be similar. Most microbes and viruses, we must note, are actually non-pathogenic (less than 1% are pathogenic!).14 Therefore, there must have been an original purpose and some change has left several of them harmful to other organisms.
Francis15 suggests a type of biomatrix between microbes and their interaction with macroorganisms. Some non-pathogenic organisms could have undergone genetic or environmental changes that led to harm. Have there been degenerative losses of good features that caused microbes to become harmful? Though possible, this is not a simple or flawless hypothesis, as pathogenic bacteria genomes are not significantly different in size than a non-pathogenic bacteria.16 If there were genetic losses that caused harm, there is not outstanding evidence for them.

Biological Similarity and Design
Creationists have always expressed great interest in biological similarity and design. This is in part because of the challenges of natural evolutionary models, but it is also because God is a good creator who designs beautiful things!17 Knowing who God is allows us to study His creation in a special way. One of those ways is appreciating similarities between organisms. Man is able to recognize patterns and features that God has placed into His creation, and this is an evidence of intentional design.
Other features of design may include mosaic-like organisms that possess both derived and primitive-looking features, as special adaptations for the environments in which they live. For example, it is suggested by creationists like Kurt Wise that Tiktaalik (traditionally thought to be a fish-to-tetrapod transitional form) was uniquely designed for a “floating forest” type of environment that may have existed in the pre-Flood world. Other “transitional forms” need more evaluation from creationist research, and this is an exciting field of study to learn about the special, unique, and exotic organisms God has made.
Creation research is a dynamic, growing, and changing field. Science is an ever-changing and developing field, and as new discoveries are made, creationists are able to refine models for origins and the Flood, as well as how organisms are related and designed. We can study God’s creation and see His attributes which bring Him glory.
“For his invisible attributes, namely, his eternal power and divine nature, have been clearly perceived, ever since the creation of the world, in the things that have been made….”
Romans 1:20 (ESV)
Footnotes
- Marsh, F.L. (1947). Evolution, Creation and Science. Washington, DC: Review and Herald Publishing Association ↩︎
- Wood, T.C. (2008a). Baraminic distance, bootstraps, and BDISTMDS. Occasional Papers of the BSG, 12, 1-17. ↩︎
- Wood, T.C. (2010). Baraminological analysis places Homo habilis, Homo rudolfensis, and Australopithecus sediba in the human holobaramin. Answers Research Journal, 3, 71-90. ↩︎
- Cavanaugh, D.P., T.C. Wood, and K.P. Wise (2003). Fossil Equidae: a monobaraminic, stratomorphic series. In R.L. Ivey (Ed.), Proceedings of the Fifth International Conference on Creationism (pp. 143-153). Pittsburgh, PA: Creation Science Fellowship. ↩︎
- Nelson, B.C. (1927). “After its Kind” The First and Last Word on Evolution. Minneapolis, MN: Augsburg Publishing House. ↩︎
- Tinkle, W.J. (1967). Heredity: A Study in Science and the Bible. Houston, TX: St. Thomas Press. ↩︎
- Lightner, J.K. (2008a). Karyotype variability within the cattle monobaramin. Answers Research Journal, 1, 77-88. ↩︎
- Wise, K.P. and M. Croxton (2003). Rafting: a post-Flood biogeographic dispersal mechanism. In R.L. Ivey, Jr. (Ed.), Proceedings of the Fifth International Conference on Creationism (pp. 465-477). Pittsburgh, PA: Creation Science Fellowship. ↩︎
- Froede, C.R. (2003). Dust storms from the sub-Saharan African continent: implications for plant and insect dispersion in the post-Flood world. Creation Research Society Quarterly, 39, 237-244. ↩︎
- Stambaugh, J. (2008). Whence cometh death? A biblical theology of physical death and natural evil. In T. Mortenson and T.H. Ury (Eds.), Coming to Grips with Genesis (pp. 373-397). Green Forest, AR: Master Books. ↩︎
- Stambaugh, J.S. (1992). “‘Life’ according to the Bible, and the scientific evidence”, Creation Ex Nihilo Technical Journal, 6 (2), 98-121. ↩︎
- Wilson, G. (2004). The origins of natural evil. Occasional Papers of the BSG, 4, 8. ↩︎
- Wilson, G. (2007). Designed for defense: reptiles and amphibians thwarting predators in a fallen world. Occasional Papers of the BSG, 10, 22-23. ↩︎
- Francis, J.W. (2003). “The organosubstrate of life: a creationist perspective of microbes and viruses,” Proceedings of the Fifth International Conference on Creationism, 433-444. ↩︎
- Francis, 2003, 433-444 (Footnote 14) ↩︎
- Wood, T.C. (2007). The Origin of Pathogenic Bacteria By Degeneration or Design, Occasional Papers of the BSG, 10, 8-9. ↩︎
- Henriksen, J. (2010). “An Architectural Paradigm“, Journal of Creation, 24 (1), 99-105. ↩︎










This article shares valid observations based not only on common-sense facts discovered through our five senses but adjusts those observations to revelation facts discerned by faith in our all-wise and all-powerful Creator and His creation of the heavens and the earth.
Congratulations to Noel Grimes for her excellent article.
Great article! There’s so much more to learn about God’s creation. I appreciate the scientists who devote their lives not only to discovery but also to the One who made all there is to be discovered.
Ame n. All research leads to the bible being true.
Yes biogeography is a creationist friend. however we must see another continent movement, up and down, to explain pathways and then ending them. In other words biology had to quickly fill the post flood world and then stop people filling it easily in order, I suggest, to fulfill the prophecy of one son , Japheth, being enlarged. the world , post flood, still was to obey Gods commands.
Great! I will be back…solid teaching!