
An intermediate form is an organism that seems to share traits with two or more different types of organisms. Archaeopteryx (between reptiles and birds), australopithecines (between humans and non-human primates) and Ambulocetus (between hoofed mammals and whales) are three popular examples. But over the past two decades, one group of intermediate forms has risen to stardom: the fishapods.
The following is a summary of “Baraminological analysis of Devonian and Carboniferous tetrapodomorphs“ by Paul Garner, and of the surrounding discussion and research pertaining to it. The views expressed are not necessarily those of New Creation.
Finding Fishapods
“Tetrapod” is a technical term applied to all forms of life based on a four-limbed body plan. Fishapods were not quite like tetrapods or fish. Much like fish they had gills and stout, fleshy pectoral fins. However, like tetrapods they also had lungs, a mobile neck, and special joints in their front limbs that functioned as elbows. Researchers find their fossils in Devonian rock layers of the geologic record. There are a number of different species, some more fish-like, and others more tetrapod-like. By far the most well-known of the fishapods is Tiktaalik, unearthed on an island northwest of Greenland called Ellesmere Island.
The unique trait combination of fishapods has led conventional scientists to suggest that they are evolutionarily related to both fish and tetrapods. They claim these serve as the transitional forms in between them. According to evolutionary theory, the origin of tetrapods from fish-like ancestors was one of the major events in the history of life.
Needless to say, this scenario doesn’t seem to fit within a young-earth history theory. According to Genesis 1:20-31, fish cannot be evolutionarily related to any land animal. God created them on separate days, fish on Day 5 and land animals on Day 6. So how can we understand fishapods from a biblical, young-earth perspective? For starters, we need to take a close look at the data and determine what we can learn from it.
A Fishy Order
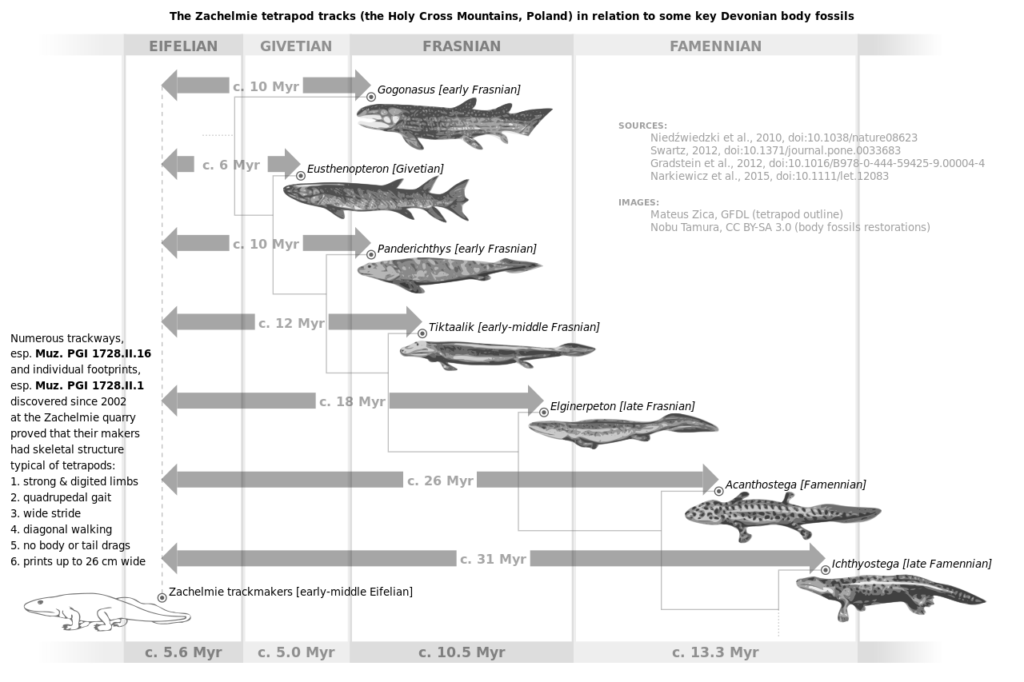
As already mentioned, we find fishapods in Devonian rock layers, but more specifically we find the various species in a very specific order. The lowest fishapods to appear in the fossil record are typically more fish-like in appearance, like Panderichthys. Further up, we find species that are more like tetrapods (amphibians to be specific). Researchers call these ichthyostegalians, represented by species such as Ichthyostega itself. Finally, in between these two extremes, we find species like Tiktaalik, Elpistostege, and other mosaics. While there are some outliers (like the Polish tetrapod tracks and the poorly-known Livoniana) the order from fish-like to more tetrapod-like species is quite consistent.
Paleontologist Kurt Wise labels fossil sequences such as this as a “stratomorphic series,” a series of fossil genera found in sediment layers, one after the other, that occur in a series of forms. We can liken this to footprints in a trackway through time. Even if evolution is not responsible for the specific appearance of various fishapods in the fossil record, it is imperative that creationists develop a model that explains their order.
Relative Ancestry
From Garner et al, 2018
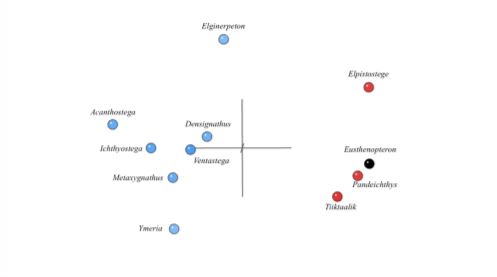
matrix with a subset of 11 taxa. Devonian tetrapods are shown in blue, elpistostegids in red and other fishes in black.”
From Garner et al, 2018
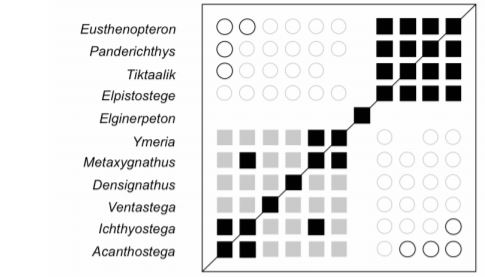
Back in 2003, creationist Paul Garner argued from his own analysis that he found Devonian tetrapods such as Ichthyostega and Acanthostega distinct enough from fishapods like Panderichthys to hypothesize that they were separated by a significant morphological (and, by inference, phylogenetic) discontinuity.1 Decades later, Garner and Jonathan Asher tested this prediction using statistical baraminology.2 In a nutshell, this is a two-step method involving baraminic distance correlation (BDC) and multidimensional scaling (MDS) that detects any morphological discontinuities that may exist between organisms.
What they found was that there was little evidence of continuity, and significant evidence of discontinuity, between fishapods and tetrapods. While it requires more to delineate the results, the findings of this study indicate that these groups of organisms originated from separate ancestry. This is very consistent with the creationist worldview, which would predict these animals belonged to different created kinds.
How Do We Explain Fishapods?
But if fishapods and tetrapods are not related via common descent, why do they consistently appear in the same fossil order wherever researchers find them? Dr. Wise incorporated the fishapod-to-tetrapod series into a theory in a paper he published in 2003.3 He suggested that the plants that have converted into coal today were the remains of a continent-sized forest that floated on the oceans of the pre-Flood world. This model, the floating forest theory, is similar to floating islands and quaking bogs of today. These unique floating forests would accommodate equally unique plants and animals, including griffinflies (seagull-sized dragonfly lookalikes), six-foot millipedes, and other large arthropods. Today, we find these organisms primarily preserved in Devonian and Carboniferous rock layers of the eastern seaboard and Europe.
An Intermediate Ecosystem
Dr. Wise points out that it is not just the fishapods that follow a particular sequence in these layers; so do the plants. And just like the fishapods, scientists interpret the plant fossil record in these layers as an evolutionary sequence. It begins horneophytes, then rhyniophytes, zosterophyllites, progymnosperms, and a botanist’s brew of herbaceous lycopods, ferns, seed ferns, and arborescent sphenophytes and lycopods.
Dr. Wise found that what both series share in common is that they are both sea-to-land sequences. In other words, as one ascends the Devonian/Carboniferous layers of eastern North America and Europe, these fossil sequences start with species that are very water dependent and are increasingly terrestrial in the successive layers further up. Dr. Wise proposes that the order of first appearance these organisms exhibit displays not the order in which they evolved, but the order in which they were overcome and buried by the catastrophe of Noah’s Flood.
According to Dr. Wise, God specially designed fishapods as morphological intermediates to inhabit an aquatic ecosystem that was itself intermediate between sea and land. He postulates that aquatic species, such as Panderichthys, lived along the outskirts of the floating forest. Tiktaalik and Elpistostege inhabited the shallow pools on the thinner portions of the floating forest, perhaps using their tetrapod-like limbs to navigate from pool to pool. Finally, aquatic creatures that needed firmer terrain from time to time, such as Ichthyostega, resided in the thicker portions of the floating forest where it could support their weight.
The Order of Destruction
Dr. Wise suggests that the forest’s destruction began near the start of the Flood. Others, like Dr. Ken Coulson, propose that there was increasing geologic activity leading up to the Flood. If so, the floating forest, designed for calm settings, may have begun to break apart even before the beginning of the Flood. Regardless, the destruction of the floating forest occurred from the outside in. The edges of the floating mat, along with the more aquatic plants and animals living there, were the first to go. They were buried on the seafloor below. Before long, the inner reaches of the forest were destroyed and sank as well. In such a scenario, the fish-to-tetrapod sequence is not evolutionary at all, but reflects pre-Flood ecology and the order of burial during the Flood.
Conclusion
Like many evolutionary theories, the fish-to-tetrapod theory makes sense of the fossil succession displayed in this group of animals. But it doesn’t account for every factor. For example, it is unable to satisfactorily explain the Polish tetrapod tracks or the discontinuities between fishapods and other organisms. Anomalies like these can be a great clue that creation researchers would benefit from further investigation here and propose models that explain the same data (and then some!) better than conventional ones.
The floating forest theory is an excellent example of such a model, but there is much more work to do. Many Devonian fish, fishapods, and tetrapods are known only from fragmentary remains. Could the discovery of new species with more complete fossils bridge the discontinuities between these groups? Or will the discontinuities identified by Garner et al be maintained? There are other stratomorphic series that await explanation within a young-earth context. What are we to make of the synapsid-to-mammal series or the ape-to-human series? Could these have something to do with the ecological order of burial during the Flood, or perhaps successions of ecological niches as the world recovers from the Flood? We’ll only know if we put in the hard work!
Footnotes
- Garner, P. 2003. “The fossil record of ‘early’ tetrapods: Evidence of a major evolutionary transition?” Journal of Creation 17(2), 111–117. ↩︎
- Garner, P.A., and J. Asher. 2018. “Baraminological analysis of Devonian and Carboniferous tetrapodomorphs,” In Proceedings of the Eighth International Conference on Creationism, ed. J.H. Whitmore, 458–471. ↩︎
- Wise, Kurt P. (2003) The Pre-Flood Floating Forest: A Study in Paleontological Pattern Recognition,” Proceedings of the International Conference on Creationism: Vol. 5, Article 31. ↩︎










There is a better creationist idea to explore. Namely that the whole classification system made by mankind is not gods classification. God made kinds. He never said he made bigger groups like mammals. reptiles, dinosaurs, or anything else. So its a option that cross creatures likeness in traits has nothing to do with divisions in nature but simply a good idea that the trait fits the creature. In limited options in biology. Why not? Creationists will get also messed up trying to squeeze biology into these invented groupings i suggest.
It’s funny you don’t make conclusions based on an unbiased read of the evidence. All creationist ever do is try to force fit the data into their misreading of the Bible. But hey that’s creationism is.
Thank you for your feedback, Jacob. If you have any specific points or counterarguments you’d like to discuss, please feel free to share them, and I’d be happy to address them in a constructive manner. This will help us better understand the nuances of the topic and promote a more informed and respectful discourse.