
By Nobu Tamura (http://spinops.blogspot.com) – Own work, CC BY 3.0, https://commons.wikimedia.org/w/index.php?curid=19459344
Whales, dolphins, and porpoises are surely among the most majestic creatures on the planet. But for as long as anyone can remember, their origin has been the subject of heated controversy. Most biologists believe that they evolved from a four-legged land ancestor (Thewissen, 2009). Young-earth creationists, on the other hand, have typically argued that whales were specifically designed as aquatic creatures (Sarfati, 2012).
The views expressed in this artile reflect those of the author mentioned, and not necessarily those of New Creation.
There has been no shortage of arguments against whale evolution from the creationist camp. Most of these tend to focus on demonstrating how certain fossil species cannot have been the evolutionary ancestors of whales, or the enormous anatomical challenges that would be required for the transition from a land mammal to a marine whale.
While this technique is quite helpful in revealing the inadequate nature of certain evolutionary ideas, it also leaves a gaping hole: how do we actually explain the evidence for whale evolution from an alternative perspective? As creationists, we should not settle for simply poking holes in evolutionary theories of whale origins. We should also attempt to explain the same data evolutionists can, but from a biblical perspective. We need to develop a framework that explains whales and their origin.
In this article, we will first search Scripture to see what is and is not permitted regarding the origin for whales. Then we will turn to the scientific data to see what we can learn from it. Finally, we will present a couple of models for whale origins and determine how they make sense of the data.
Denizens of the Deep

Whales, dolphins, and porpoises belong to a group of animals called cetaceans. Despite living in the water, they are not fish; they are mammals, just like dogs, elephants, mice and kangaroos. There are certain features that scientists use to distinguish mammals from other animals. They must be 1) warm-blooded, 2) breathe air, 3) give birth to live young, 4) nurse their young with milk and 5) have hair (Thewissen, 2009). Modern whales swim with the aid of their muscular tail and forked tail fluke that moves up and down to propel them forward. Cetaceans can be divided up into three main groups.

First are the mysticetes, or baleen whales. This group contains the largest animals that have ever existed, like the blue whale and the humpback whale. Blue whales can grow over 98 feet long (Sears, 2002) and weigh more than 173 tons. (McClain, 2015) Instead of teeth, these whales have rows of long plates in their mouths covered in bristle-like structures. These are called baleen plates, and the whales use them to filter small animals, mostly krill, from the surrounding water.

Second, we have the odontocetes, or toothed whales. This group contains dolphins, porpoises, orcas, sperm whales, narwhals, and belugas. The sperm whale is the largest, measuring up to 66 feet from snout to tail fluke and weighing more than 45 tons. (McClain, 2015) Toothed whales are well-known for their ability to echolocate, allowing them to locate food, potential threats, and other whales, even when visibility is low.

By Ghedoghedo – Own work, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=16288308
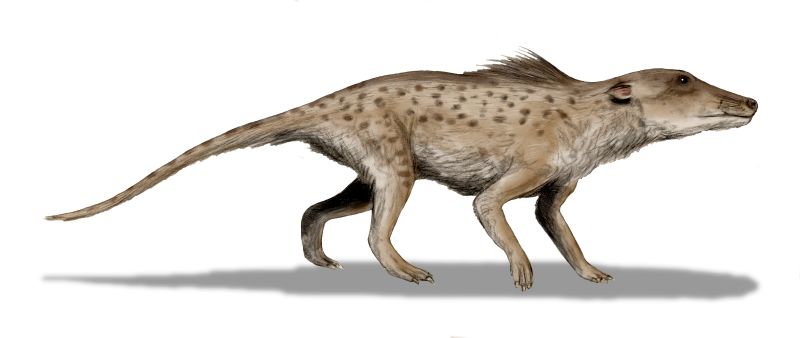
Finally, there are the archaeocetes, or ancient whales. They came in far more varied forms than the other two groups. Some, like Dorudon and Basilosaurus, are fully aquatic like the whales of today. Others, such as Maiacetus, are semi-aquatic, almost walrus-like creatures that spent much time in the water, but occasionally returned to land to rest and give birth. And still there are a number of archaeocetes, like Pakicetus, which are semi-terrestrial, somewhat like hippos are today. Archaeocetes are only known from the fossil record, with no living representatives in the present.
Groups within Groups
You may be wondering how animals as different as the blue whale and Pakicetus can both be cetaceans. On a surface level, this has nothing to do with evolutionary relationships, and everything to do with the way we classify life. The classification of living things into a groups-within-groups pattern is called taxonomy. It makes it easier for scientists to easily communicate with colleagues about what specific type of organism is being studied and why. Instead of saying, “I study animals that have fur, give birth to live young which they nurse with milk, and are warm-blooded,” a scientist has a much easier time of it stating, “I study mammals.”

The common names we give to groups of living things in everyday speech are often different from those used by scientists. For example, a layperson would define a whale as a large marine mammal with a fluked tail and a blowhole. The colloquial names we give to animals often reflect their attributes (e.g. sawfish) or behaviors (e.g. woodpecker). According to the Online Etymology Dictionary, the word ‘whale’ can be traced back to a Proto-Indo-European the word *(s)kwal-o-, which means “large sea fish.” This makes sense, given that the term “fish” was a more inclusive term at that time and used to refer to a variety of water-dwelling animals.
The technical, or scientific, name given to animals by specialists reflects which group they belong to in respect to the entire Tree of Life. Scientists distinguish different groups of animals from each other based on one or more characteristics. Cetaceans are set apart from other mammals by the unique shape of their inner ear bones (Thewissen, 2009). The inner ear is an air-filled cavity in the skull. The walls surrounding this cavity in most mammals are generally the same thickness all the way around, but not in whales! The internal wall of their inner ear cavity is thicker than the external wall surrounding it. Scientists call this thickened wall the involucrum, and it is something all whales have. No non-cetaceans have it except a small extinct mammal called Indohyus. Because they share this feature with true whales, some scientists think Indohyus should also be classified as a cetacean, (Thewissen, 2009) but this is up for debate and beyond the scope of this article.
The fact that all cetaceans possess an involucrum does not automatically mean they are related via common descent. This is an evolutionary assumption. Because evolutionists believe that all biodiversity arose by purely natural processes, any similarity shared between two organisms must be interpreted as deriving from a common ancestor except for when there is evidence to the contrary. On the other hand, creationists do attribute some similarities to common descent, but they also expect other similarities to have resulted from a common design or convergence.
What Does the Bible Have to Say?
Before we go any further, it is of utmost importance that we wander not from our foundation. As biblical creationists, we need to test our understanding of the created world against the Creator’s Word. Let’s examine what it does and doesn’t say about the origin of animals.
“And God said, ‘Let the waters swarm with swarms of living creatures, and let birds fly above the earth across the expanse of the heavens.’ So God created the great sea creatures and every living creature that moves, with which the waters swarm, according to their kinds, and every winged bird according to its kind. And God saw that it was good…And God said, ‘Let the earth bring forth living creatures according to their kinds—livestock and creeping things and beasts of the earth according to their kinds.’ And it was so. And God made the beasts of the earth according to their kinds and the livestock according to their kinds, and everything that creeps on the ground according to its kind. And God saw that it was good.”
Genesis 1:20-24
From this passage, we learn that God created the animals according to their kinds. He created various kinds of water animals on Day 5 of Creation Week, and various kinds of land animals on Day 6. Whales are water animals, so they must have been created on Day 5 as fully-aquatic animals from the beginning. Case closed…right? Not quite.
While it’s true that all species of whales today live in the water, we must remember that nowhere in Scripture does it say that species cannot change. In fact, the word “species” is never used anywhere in the original Hebrew text! It instead uses the word “kind.” A biblical “kind” can be thought of as an original, interfertile population of organisms God created, as well as their descendants. Since there are many different meanings for the word “kind” in the English language, we will proceed with another, more specific term: “baramin,” which is Hebrew for “created kind.”
There can be many species, genera, subfamilies, and sometimes even families within a baramin (Lightner, 2012). For example, lions, tigers, house cats, and saber-toothed cats all belong to the cat baramin because they descended from the ancestral cat-like creatures God created on Day 6 (Pendragon, 2011).

By MathKnight – Own work based obגירית מצויהגרגרןגירית הדבשהרמיןלוטרה אירופיתדלק אירופי, CC BY-SA 4.0, https://commons.wikimedia.org/w/index.php?curid=86133534
Notice, though, that the Creation account in Genesis says nothing of what the limits of diversity are within each baramin. Even without accepting a land-based origin for whales, we can still appreciate the fact that the members of a baramin have great capacity to change, even to the point of adopting a completely different lifestyle. The kakapo, for instance, is a member of the parrot baramin from New Zealand that lost the ability to fly (Lightner, 2013). It fills much the same ecological niche as a small mammal, even to the point of becoming nocturnal and developing the keenest sense of smell among the parrots (Hagelin, 2004). The sea otter is another example. Despite likely coming from the same common ancestor as weasels, badgers, and wolverines (Lightner, 2012), these playful critters have developed adaptations essential to becoming a marine mammal, including flippers. However, individuals belonging to one baramin are unable to change into completely different baramins (i.e. ape-like creatures into humans).
But wait! Doesn’t Genesis specifically state that God created whales on Day 5? Some Bible versions, like the King James, do in Genesis 1:21. However, the original Hebrew word translated as “whales” here is תַּנִּין, or tannin. This word has no known parallel in modern Hebrew and thus has proven quite difficult to translate. However, it has often been translated elsewhere in the King James Version as “dragon,” “great sea creatures” in the English Standard Version, and “reptile” in the Septuagint text. Tannin is used again in Exodus 7:9-12 to refer to the creature Aaron’s staff became: a serpent. Genesis 1:21 could easily refer to any number of large water-going creatures and not specifically whales.
Walking with Whales
Having established that a land-based origin for whales is, at the very least, scripturally permissible, we can now examine the scientific evidence for this claim.
Modern whales have two external limbs called flippers, but they also have a pair of pelvic bones inside their bodies where the hip is located in land animals. Useless for walking or swimming, they are used to support the reproductive organs. Evolutionists cite these as vestigial structures, body parts that have lost their original function and have been reduced as a result.
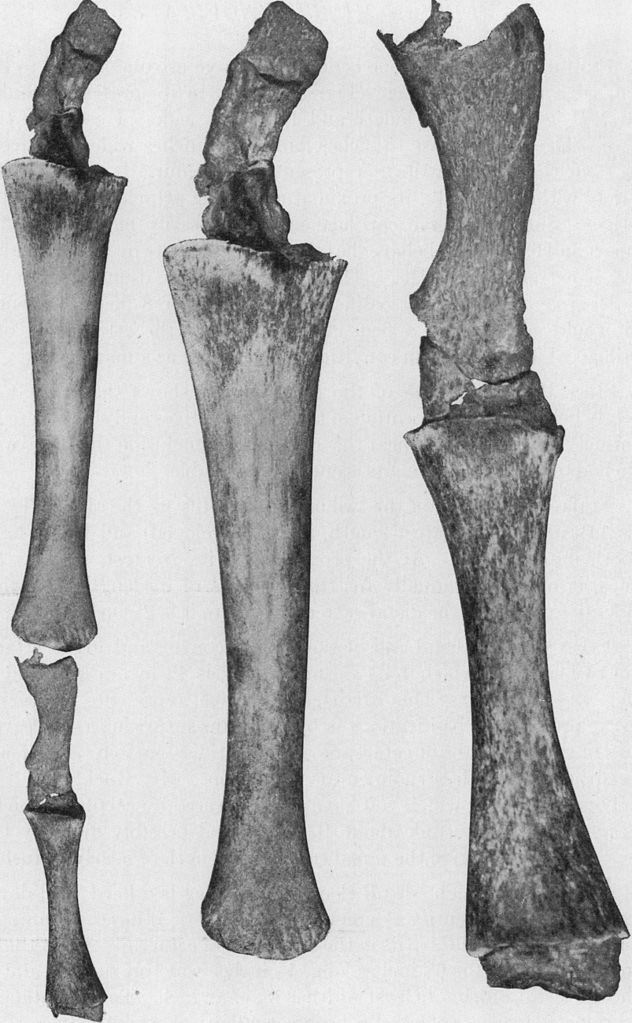
Roy Chapman Andrews, Public domain, via Wikimedia Commons
Interestingly, whales develop external hind limb buds whilst in their mother’s womb. These usually shrivel up and disappear before the whale calf is born, but on rare occasions they are retained into adulthood. One such discovery was of a female humpback whale that occurred in July of 1919 (Andrews, 1921). Its remains were analyzed by renowned American Museum of Natural History paleontologist Roy Chapman Andrews. The 45-foot marine mammal had two back limbs that were 4.2 feet long and composed of femur and thigh bones, tarsal and metatarsal bones, and even digits. The limbs ended in a rounded knob that was shaped like a man’s fist. More recently, on October 28, 2006, a bottlenose dolphin was accidentally captured by fishermen off the coast of western Japan (Thewissen, 2008). It had two hind limbs at the base of its tail. Evolutionists commonly cite instances like these as examples of genetic throwback, where an organism is born displaying a character trait derived from an ancestor.
Turning to the fossil record, we find a plethora of back-limbed whales: the archaeocetes. Basilosaurus and Dorudon have the overall body plan of modern whales, but also tiny hind limbs. And of course, “walking whales” like Pakicetus come in no short supply, and they look very much like a four-legged version of Dorudon.

By Ideonexus – [1], CC BY-SA 2.0, https://commons.wikimedia.org/w/index.php?curid=6053706
It isn’t just that a wide variety of whales have been found, but they are also said to occur in a particular order called a stratomorphic series. This is when a group of fossil organisms is arranged into successive rock layers, forming a gradient from one form to the next. You may be familiar with the famous fossil horse series.
Everything listed above is used as evidence that whales evolved from a four-legged cetacean that lived on land, and that they share a common ancestor with other mammals. Recall, though, that this is based on the premise that similarities shared between two or more organisms generally must have been derived from a common ancestor. What if we adopt the young-earth creationist perspective and do not take this assumption for granted? How might we interpret this evidence based on the premise that God created the animals according to their kinds?
Four options present themselves:
- The fossil whale series is nothing but a figment of the evolutionists’ imagination. In other words, the evolutionary diagrams of whale evolution do not accurately reflect the order in which the various types of cetaceans appear in the fossil record. If this is the case, then the similarities we observe between different types of cetaceans can be attributed to common design or convergence.
- The fossil record of cetaceans represents the order in which different cetaceans were buried, either during the Flood or perhaps reflecting where they lived after the Flood.
- The fossil record of cetaceans simply represents their diversification from a pair of legged archaeocetes that walked off the Ark after the Flood.
- A combination of two or more of the previous options.
The Sweetest Series?
“During the past fifteen years, new discoveries in Africa and Pakistan have greatly added to our paleontological knowledge of the earliest history of whales. The embarrassment of past absence has been replaced by a bounty of new evidence—and by the sweetest series of transitional fossils an evolutionist could ever hope to find.”
– Stephen Jay Gould, “Hooking Leviathan by its Past”
As stated previously, there is an alleged series of fossil whales, starting with Pakicetus in the lowest layers, four-limbed whales like Rodhocetus and Dorudon somewhere in the middle, and modern two-limbed whales in the highest layers. No doubt you’ve seen the whale series portrayed in diagrams before. But is it valid?
Though we disagree with the million-year estimates assigned to fossils, most young-earth paleontologists readily accept that fossils in general are found in a specific sequence that is consistent anywhere you go on the earth. The order in which fossils are found usually correlates to their order of burial (i.e. fossils in lower rock layers were buried prior to fossils in higher rock layers). Zooming in on whales, we see that some very interesting patterns begin to emerge.
In a general sense, the various cetaceans in the fossil record occur in a consistent order (Thewissen, 2009). The first four-legged cetaceans tend to appear in rock layers lower than the fully-aquatic basilosaurids, which are then followed in higher rock layers by the appearance of toothed whales and baleen whales.
However, it is in the specific sequence of fossil cetaceans where things begin to break down. Conventional paleontologists have given Pakicetus an age range of approximately 56 to 41.3 million years ago, while Ambulocetus, Remingtonocetus and Rodhocetus have all been given an age range of 47.8 to 41.3 million years ago. (Given that Ambulocetus is known from only one skeleton, it is impossible to determine its absolute age range; nevertheless, the available data fits well with the previously mentioned archaeocetes.) From this, it seems that many of the legged archeocetes were actually contemporaries of each other and may not represent an evolutionary lineage after all.
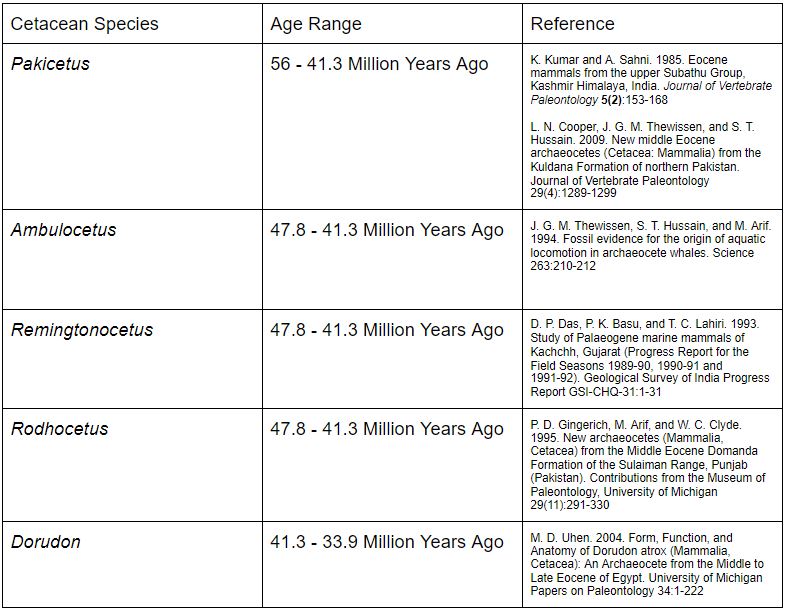
It seems that the hypothesis that the whale fossil series is a figment of evolutionary imagination is without grounds and can be rejected. However, the fact that the order is nonspecific in many cases is an indication that a non-evolutionary explanation for these fossils may be applicable here.
Order of Burial
Because organisms must usually be buried quickly in order to be preserved, it might be tempting to argue that the fossil whale series was the product of burial order during the Flood.
One of the problems with this is that most young-earth paleontologists do not in fact believe that any whale fossils are from the Flood. Whales are found exclusively in the uppermost portion of the fossil record, called the Cenozoic. This is largely thought to represent successive snapshots of ecological recovery as early post-Flood animals and plants were buried by local or regional-scale catastrophes.1
Another problem is that the whale fossil series does not fit what would be expected from the Flood. One major factor young-earth paleontologists think contributed to the Flood’s fossil record is the order in which pre-Flood ecological communities of plants and animals were overcome by the rising floodwaters. This would produce a sea-to-land sequence. Unfortunately, the whale fossil series is the exact opposite: it is a land-to-sea sequence (Wise, 1995).
What post-Flood factors may have contributed to the fossil whale series? A helpful analogy may come from another stratomorphic series: the fishapods. Hailing from the Devonian geologic system, the fossils in this series are found in a particular order. First to appear are aquatic fish, such as Panderichthys, followed further up by tetrapods with true legs such as Acanthostega, and the intermediate fishapods, like Tiktaalik, show up in the layers between them. Though usually interpreted as an evolutionary sequence, young-earth paleontologists suggest that fishapods had intermediate morphology because they lived in an environment that was itself intermediate between sea and land. The aquatic fish, fishapods, and tetrapods of the Devonian lived at the same time, but in adjacent ecological zones.
How might we apply this to whales? First off, Cetacea may be polybaraminic. While a baramin is a group of organisms belonging to the same created kind, a polybaramin is a group of organisms belonging to different created kinds. So if Cetacea is polybaraminic, that means it would be composed of more than one created kind.

In this scenario, semi-terrestrial and semi-aquatic archaeocetes (e.g. Pakicetus, Ambulocetus, Rodhocetus) could belong to one or more baramins that came off the Ark and diversified after the Flood. Meanwhile the fully-aquatic archaeocetes (i.e. Basilosaurus), toothed whales, and baleen whales could represent one or more baramins that survived the Flood, but for one reason or another are absent from the fossil record until some time afterward. These whales would also diversify in the post-Flood world. The fossil whale series, similar to the fishapod series, may represent a gradient of land-to-deep sea whale baramins living in a gradient of land-to-deep sea environments (ecozones, if you will). This may serve as a better explanation for the fossil record of legged archaeocetes, given that they seem to have been contemporaries of each other.
Also supporting this hypothesis is the fact that archaeocetes are almost always found in shallow water sediments (Wise, 1995). It may be that fossilization in the lower Cenozoic (earliest post-Flood era) was more likely in terrestrial and shallow water environments where the semi-aquatic archaeocetes were living. Or maybe, having been reduced in number by the Flood, fully-aquatic whales did not become widespread or common enough to leave fossil evidence behind until the post-Flood era was well underway. Alternatively, what if there has been a bias towards searching shallow-water fossil deposits versus fossil deposits reflective of deep water environments (Wise, 1995)? All of these are viable hypotheses that should be put up for consideration for researchers with interest in this field.
Taking to the Water

By Allosaur – Создал сам, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=7875453
Another possible explanation for the fossil whale series is that it truly does represent the diversification of one whale baramin after the Flood, similar to what creation researchers have proposed concerning the fossil horse series. This may seem an odd thing to discuss within young-earth creationist circles, but there are surprisingly quite a few individuals willing to give this idea consideration.
Paleontologist Kurt Wise is probably the most well-known creationist who is bearing this idea in mind (Wise, 2009), but he is not the only one. Creation researcher Colin Brown has also spoken about the plausibility of this theory (Brown, 2010). Some advocates of Recolonization Theory suggest that not only did whales make the transition from land to sea, but manatees and dugongs, ichthyosaurs, and sea turtles did as well.
Some creationists have accused Dr. Wise of being a “hyper-evolutionist” or a “young-earth evolutionist” (Hebert, 2020) These critics argue that he is simply accommodating “absurd evolutionary claims at face value.” This is untrue. In reality, this is simply Dr. Wise’s default position. As of yet, he is unconvinced that Cetacea is composed of more than one baramin until suitable evidence for an alternative is brought to the table (Wise, 2021).
Currently, Dr. Wise tentatively suggests that when God made the first cetaceans during Creation Week, they were semi-terrestrial or semi-aquatic creatures, perhaps similar to Rodhocetus. After the Flood, a pair of Rodhocetus-like cetaceans walked off the Ark and multiplied. As their descendants diversified, they gave rise to the rest of the whale baramin. Some of these descendants took to the water, filling the empty niches left vacant by the extinction of marine reptiles during the Flood.
Assuming for a moment that whales did take to the water sometime after the Flood, how might they have gone about doing so, biologically speaking, and in the time allotted us from Scripture? This is not just a mystery we must solve for whales, but for all modern biodiversity. Even if we do not accept a land-based origin for whales, creationists must still account for the great variety of life apparent today that descended from just a few thousand baramins on the Ark. Recall that most modern species were in existence by Abraham’s time, somewhere between a couple of centuries and a thousand years after the Flood (Wood, 2002). Such grand biodiversity is not achievable by conventional diversification methods. The key word being conventional.
See, most naturalistic evolutionists credit the diversity of life largely to natural selection acting upon variation potential that arises from random mutations. This process is painfully slow. It would take many millions of years to generate post-Flood biodiversity in this fashion.

By Huangdan2060 – Own work, CC BY 3.0, https://commons.wikimedia.org/w/index.php?curid=87383026
But creationists aren’t hampered by the snail-paced rate of change required by Darwinian evolution. We do acknowledge the reality of mutations and natural selection, but we are also beginning to realize that other diversification mechanisms may have been involved. While there is still much debate as to what mechanisms these were, there is general agreement that it had something to do with created heterozygosity. In other words, biodiversity within a baramin was made manifest based upon the great variation potential already encoded in their genes upon creation. As the baramins left the Ark and spread across the earth, their descendants readily adapted in order to colonize vacant ecological niches. Novel morphologies and behaviors were generated in response to specific environmental pressures. Interestingly, the Cenozoic radiation of cetaceans is quite fast by naturalistic evolutionary standards as well; the oldest fully aquatic whales, like Basilosaurus, appear in the fossil record within ten million years or so after the first Pakicetus-type creatures (Thewissen, 2009) on the conventional geologic timescale.
Colin Brown summarizes what such a land-to-sea transformation for whales might look like from a creationist perspective:
“[L]et us look for a moment at what prospects there are for producing a basic whale from a land creature, because if this were possible our options as creationists with respect to the whale would be far greater and would silence evolutionists on this subject altogether! If we are comfortable with seals originating on land, producing many inches of blubber, and having limbs transformed into flippers, then we have only to take this a little further and we could have the dugong (Dugong dugong), which does not have any hind limbs but does have a tail fluke like the whale. From here we only need to have the head become elongated, and there is nothing wrong with this in creation, for things can get larger/smaller/fatter/thinner etc. in creation; no laws are broken by this. Nasal openings then move from the snout to the top of the head, and we have our first basic whale, from which subsequent changes could arise. There is a creature in the fossil record with nasal openings about midway between snout and the top of the head, namely the Basilosaur. This could possibly happen because the [baramin], to me, is a plan that includes both what we can see, i.e. the phenotype, and what I call the life-form’s genetic potential. This is the amount of genetic change possible, taken from any given point in time into the future.”
Brown, 2010
Similarly, Dr. Wise states that the land-to-sea transition in whales might make more sense in light of the young-earth paradigm:
“[T]he ongoing process of mutation destroys genetic information. Although natural selection does tend to eliminate expressed harmful mutations (as less fit expressions of a gene), natural selection cannot eliminate unexpressed mutations. Since the average nucleotide will be mutated every million generations or so, a million generations should alter every single nucleotide of a gene, rendering it completely unusable—even unrecognizable as a gene. In a few million years, there should be absolutely no genetic information left in unused genes. Vestigial organs and genetic throwbacks suggest that the genetic information has not been unused for very long, and if modern whales are descendant from the legged whales in the fossil record and modern horses are descendant from the three-toed horses in the fossil record, then this has happened recently—probably since the Flood only several thousand years ago. This, in turn, would suggest that the Cenozoic fossil record is not recording millions of years of time, but only thousands of years. This, in turn, would suggest that organisms diversified much too rapidly to be explained by any theory of speciation currently proposed by biological evolution. Rapid revelation of hidden species designs is the only reasonable explanation for such rapid speciation.”
Wise, 2018a
According to Their Kinds

By Cangadoba – Own work, CC BY-SA 4.0, https://commons.wikimedia.org/w/index.php?curid=72161549
We have before us two, biblically-consistent, and testable models concerning the fossil record of whales: one suggesting that all whales may belong to a single baramin, and another suggesting that the fossil record of whales represents many whale baramins buried based on their post-Flood ecology. How might we go about determining which one is the best explanation? For this, we will have to turn to baraminology.
Baraminology is the study of created kinds. This field strives to determine how many baramins there are and how they diversified since Creation and the Flood. The baraminology of whales is an underdeveloped field, but has revealed interesting information for us.
Baraminologists believe that baramins can be distinguished by identifying the discontinuity that surrounds them. One method used to help determine this is called statistical baraminology. So far, such analyses have been able to identify horses, various dinosaurs, and fishapods, among many other groups of animals and plants (Wood, 2016), as tentative baramins. What would they make of whales?

By Paleocolour – Own work, CC0, https://commons.wikimedia.org/w/index.php?curid=78254769
Biologist Todd Wood has by far done the most exhaustive work on the statistical side of whale baraminology. Starting in the mid-2000’s, he cited evidence for at least five whale baramins: 1) baleen whales, 2) beaked whales, 3) sperm whales, 4) the remaining toothed whales, and 5) the archaeocetes (Mace et al. 2005; Wood, 2006; Wood, 2007). The possibility that all archaeocetes may belong to a single baramin was particularly surprising, especially given the diverse array of semi-terrestrial, semi-aquatic, and fully-aquatic species it contains. Taken together, Dr. Wood’s research supports the hypothesis that Cetacea is a polybaramin.
Dr. Wood revisited baleen whales for the first time in over a decade and identified discontinuity within this group (Wood, 2020). He tentatively established three additional families of baleen whales―Eomysticetidae, Aetiocetidae, and Balaenidae―as baramins. The results also indicated Mammalodontidae, another family of baleen whales, may be its own baramin as well, but Dr. Wood couldn’t distinguish them with certainty from baleen whales belonging to the family Llanocetidae. If the results of Dr. Wood’s research turns out to be correct in the long run, it might lend support to the idea that much of the fossil whale series reflects the ecology of early post-Flood whales.
On the other hand, some baraminologists are dissatisfied with the evidence Dr. Wood has presented so far that Cetacea is a polybaramin. It has been pointed out that whales of any kind are completely absent from the Flood fossil record (Wise, 1995). Considering most of the Flood’s fossil record is composed of marine animals (Wise, 1988), this would be unexpected if marine whales existed at the time of the Flood. In fact, no marine mammals of any kind appear in the fossil record until well after the Flood (Wise, 2009).

By Dawn Pedersen – https://www.flickr.com/photos/paintedbydawn/34140887626/, CC BY 2.0, https://commons.wikimedia.org/w/index.php?curid=71727986
Dr. Wise also points out that many of the tentative baramins Dr. Wood has identified in his research appear in a certain order in the fossil record (Wise, 2020). For instance, deep sea trawling efforts have dredged up numerous fossils of beaked whales and other creatures from between 100 and 1,000 meters below the shore of Africa’s southern coast (Bianucci, 2008). These whales date back to the Miocene, meaning they are from the post-Flood era. Due to their size and durability, the bones from these whales have proven to be quite resilient and capable of lasting considerable amounts of time on the ocean bottom in favorable conditions. If beaked whales truly their own baramin, as Dr. Wood’s research suggests, and their bones so durable, then why do their fossils not appear until the Miocene? Dr. Wise points out that if each of the major groups of whales identified in Dr. Wood’s study represents its own baramin, then after the Flood, each one would appear for a brief period of time just as the one preceding it disappeared, only for itself to disappear as another baramin abruptly appears (Wise, 2020). This pattern would be very unusual if multiple baramins of whales were already in existence before the Flood. This is most consistent with the idea that all members of Cetacea belong to a single baramin.
An important takeaway here is that neither side holds their position dogmatically. They both understand that the field of whale baraminology is very young, there is still a lot we don’t know, and new discoveries could sway the burden of proof in either direction.
Dr. Wood sums it up this way:
“There are many whale fossils with varying sizes of hind feet, including Ambulocetus, Rodhocetus, and Dorudon. These are not modern-type toothed or baleen whales, however. They’re classified as Archaeoceti, and my research has shown that they are not the same created kind as the modern whales. Dr. Wise thinks we should keep an open mind, though, because it’s at least conceivable that all whales descend from a common ancestor. That would certainly explain the toothed baleen whales (no, I’m not kidding). My research so far does not support that, but things could change. Is this dangerous? No. It’s not. What’s dangerous is not talking about these hard problems or pretending like they’re easily solved.”
Wood, 2018
Conclusion

The acceptance of a legitimate fossil whale series or even the thought of Adam and Eve frolicking with “walking whales” in the Garden of Eden are certainly not conventional among young-earth creationists! Such imagery might even make us queasy just to think about. But creation biology is an actual science, which means it is subject to change.
Change is nothing new to young-earth creationism. Creation scientists are constantly learning both from God’s Word and the universe to develop testable scientific models that we hope will help us better understand the world around us. Sometimes, new understandings involve the alteration or rejection of scientific models we’ve developed or grown up with. The vapor canopy model is a wonderful example of a scientific model developed by creation scientists. Though popular in the mid- to late-20th century, it has since been discarded as biblically-unnecessary and scientifically-unfeasible.
Of course, this is not to say that we reinterpret the Bible to conform to the popular scientific models of the day. We need to do the opposite. Scientific deductions are only as good as the assumptions we use to arrive at them. They are subject to groupthink and insufficient data, which means that they can only ever be considered “tentative truths” (Wise, 2018). As creationists, we must interpret the world around us through the worldview lenses of God’s Word, which is the ultimate truth. What God explicitly tells us in His Word is non-negotiable. If the history of science and religion have taught us anything, it is that giving popular scientific models of the day authority over God’s Word can end in disaster. If the Bible is too tightly wound up with once-popular, but ultimately wrong notions (i.e. species fixity), many will mistakenly think that the Bible itself was debunked.
However, there is much the Bible doesn’t tell us. How did we get modern biodiversity? Was the fossil record formed before, during, or after the Flood…or a combination of all three? Did whales descend from walking cetaceans, or were they always of the sea? When Scripture is truly silent, it is then that we can use science to fill the gaps in our knowledge. When performing science from a biblical worldview, we can learn new and fascinating things about the wonderful world God created.

By Serguei S. Dukachev, CC BY-SA 3.0 http://creativecommons.org/licenses/by-sa/3.0/, via Wikimedia Commons
Acknowledgements
The author wishes to thank Dr. Matt McLain for his help in improving this article, as well as Dr. Marcus Ross and Dr. Kurt Wise for their insights into cetacean baraminology.
Notes:
1 “Tomkins and Clarey (2019) attempted to use Cenozoic whale fossils to contend for a late post-Flood boundary, though nothing in their results actually rules out an earlier boundary (particularly as their mapping emphasizes coastal fossilization within a post-Flood continental landscape).” Arment, C. (2020). To the Ark, and Back Again? Using the Marsupial Fossil Record to Investigate the Post-Flood Boundary. Answers Research Journal,13, 1-22.
References:
Andrews, R. C. (1921). A remarkable case of external hind limbs in a humpback whale. American Museum novitates.
Bianucci, Giovanni & Post, Klaas & Lambert, Olivier. (2008). Beaked whale mysteries revealed by seafloor fossils trawled off South Africa. South African Journal of Science. 104. 140-142.
Brown, C. 2010. “The Origin of Whales.” CRSQ 47(2):161. CELD ID 23968
Hagelin, Julie C. (January 2004). “Observations on the olfactory ability of the kakapo Strigops habroptilus, the critically endangered parrot of New Zealand“. Ibis. 146 (1): 161–164.
Lightner, Jean. (2012). Mammalian Ark Kinds. Answers Research Journal. 5. 151-204.
Lightner, Jean. (2013). An Initial Estimate of Avian Ark Kinds. Answers Research Journal. 6. 409-466.
McClain, C. R.; Balk, M. A.; Benfield, M. C.; Branch, T. A.; Chen, C.; Cosgrove, J.; Dove, A. D. M.; Helm, R. R.; Hochberg, F. G.; Gaskins, L. C.; Lee, F. B.; Marshall, A.; McMurray, S. E.; Schanche, C.; Stone, S. N. (2015). “Sizing ocean giants: patterns of intraspecific size variation in marine megafauna”. PeerJ. e715: e715.
Online Etymology Dictionary. “whale (n.)”. Retrieved 18 March 2018.
Pendragon, B., & Winkler, N. (2011). The family of cats—delineation of the feline basic type. Journal of Creation, 25(2), 118-124.
Sarfati, J. (2012). Chapter 5: Whale Evolution? In Refuting Evolution: A Handbook for Students, Parents, and Teachers Countering the Latest Arguments for Evolution (5th ed.). Creation Book.
Sears, R.; Calambokidis, J. (2002). Update COSEWIC status report on the blue whale (Balaenoptera musculus) in Canada. Ottawa, ON: Committee on the Status of Endangered Wildlife in Canada. p. 32.
Thewissen, J G M & Cooper, Lisa & George, John & Bajpai, Sunil. (2009). From Land to Water: the Origin of Whales, Dolphins, and Porpoises. Evolution Education and Outreach. 2. 272-288. 10.1007/s12052-009-0135-2.
Wise K. P. 2009. Mammal kinds: how many were on the ark? in: Wood T. C. and Garner P. A. (editors). Genesis Kinds: Creationism and the Origin of Species. CORE Issues in Creation No. 5, pp.129-161.
Jake Hebert, Ph.D. 2020. Is Creation Evidence Ambiguous?. Acts & Facts. 49 (8).
Mace, S.R. and T.C. Wood. 2005. Statistical evidence for five whale holobaramins (Mammalia: Cetacea). Occasional Papers of the BSG 5:15.
Wise, K.P., ‘The Flood and the fossil record’, an informal talk given at the Institute for Creation Research, San Diego (USA) on August 17, 1988.
Wise, K. P., 1995. Towards a creationist understanding of ‘transitional forms’. CEN Tech. J., 9(2):216-222.
Wise, K. Personal Correspondence. 2020.
Wise, K. Personal Correspondence. 2021.
Wise, K. P. (2018a). Perspectives on Speciation. In Devotional Biology: Learning to Worship the Creator of Organisms (pp. 352-353). Nashville, Tennessee: Compass Classroom.
Wise, K. P. (2018b). The Nature of Science. In Devotional Biology: Learning to Worship the Creator of Organisms (pp. 28). Nashville, Tennessee: Compass Classroom.
Wood TC. 2002. The ageing process: rapid post-flood intrabaraminic diversification caused by altruistic genetic elements (AGES). Origins 54:5-34. (grisda.org)
Wood, T.C. 2006. Exploration of biological character space surrounding living and fossil whales (Mammalia: Cetacea). Occasional Papers of the BSG 8:10-11.
Wood, T.C. 2007. Evidence that some toothed mysticetes are archaeocetes (Mammalia:Cetacea). Occasional Papers of the BSG, 10, 23-24
Wood, T. 2016. A list and bibliography of identified baramins. Journal of Creation Theology and Science Series B: Life Sciences 6:91-101.
Wood, T. C. (2018, July 30). Continuing the baraminology panel discussion [Web log post]. Retrieved March 29, 2021, from http://toddcwood.blogspot.com/2018/07/continuing-baraminology-panel-discussion.html
Wood, T. C., Fears, G., & Doran, N. A. (2020). A Baraminological Analysis of Fossil Mysticetes. Journal of Creation Theology and Science Series B: Life Sciences, 10, 7-8. doi:https://www.coresci.org/jcts/index.php/jctsb/issue/view/26
The views expressed in this article reflect those of the author.










Hi Ryan,
Thanks for putting together this article. I wonder if New Creation would consider publishing a response to this article which would be presented as a alternative viewpoint?
To get an idea of what this would be like you could have a look at what I have written on my website:
http://www.paradigmshift.org.uk/Whales.htm
Regards,
Marc
Thank you for this suggestion, Dr. Surtees! We will put it under consideration. 🙂
It would be great if there was a spoken version of this article so I wouldn’t have to read it to obtain the information.
That is a good suggestion, Rob! We will keep that in mind for the future.
Though a YEC critic I have to say, this was one of the best written, well-researched, informative and thought-provoking creationist’ articles I have read in a while.
Thank you, Dr. Duff! We appreciate the compliment!
I have broiught it up on your own forum but it was a great article and great to finally hopefully see a end of YEC saying marine mammals were created on creation week.Now finally they were just crusing with noah.
This was a great article by the author and a great subject. As a YEC I have always insiisted marine mammals are only post flood/ on the ark land lovers who took to the newly empty seas after the flood. its clear and demanding to see whales etc as land creatures. i also like the idea of spectrums within a kind accounting for all diversity in these creatures whether swimming or just tipping thier toes in the water or shy . Actually i would see creation week kinds as very few and even whales as just members of a lost of creatuyres. indeed hippos probably are relatives. Marine mammals are amongst the few creatures that truly show a previous bodyplan of note. otherwise biology is silent. So walking whales walks over evolutionism terribly on the very point they love to press. whales anatomical remnants of a land life make a creationist case and hurt the evolutionist case if one thinks about it.
YES creationists everywhere no more God created whales. nope. they are simply types in some critter off the ark filling the post flood earth.
Anyone with the gifts of holy spirit as listed in 1 Corinthians 12:8 would be able to discern that the firmament is Earth’s crust and the waters below are the boiling hot water that Enoch mentioned and which made the dew as Earth didn’t have rain until the Flood and after.
The waters above are obviously the vapour layer which prevented harmful sunlight burning Eve’s delicate skin.
I concur with the late paleontologist Gunter Bechly that unless the waiting time problem for the origin and spread of the required genetic changes is accounted for, the whales aren’t walking anywhere.