
With all the controversy surrounding Darwin’s On the Origin of Species, it is often easy to forget what exactly Darwin observed and wrote about after his voyage on the HMS Beagle. It is also commonplace for creationists to forget that Darwin’s book focused on the origin of species more than the molecule-to-man evolution we often associate with Darwin’s work. While he certainly did hypothesize that all living things originate from one or a few common ancestors (an idea called “universal common ancestry”), the evidence Darwin presented in his book summarizes the case for a new species arising from an ancestral species in response to selective pressures. In other words, Darwin’s core argument concerned the process of speciation more than the idea of universal common ancestry. These are two very different ideas. The former describes getting two or more species from one ancestral species, while the latter involves getting all living species from an ancient, primitive ancestor.

Over the years, Darwin’s critics have come to associate macroevolution with universal common ancestry. Perhaps without realizing it, they picked and chose what they liked about the theory of evolution (speciation) and staunchly rejected the leftovers (“macroevolution” and “common ancestry”). The result of this picking and choosing is a mantra we in creation biology have heard over and over again. Say it with me: “Microevolution good, Macroevolution bad.” In other words, small changes and variations do happen in nature, but they do not lead to the significant changes needed to turn a microbe into a monkey no matter how many billions of years you have. This reasonable position has been accepted by creationists for decades.
Is Microevolution Enough?
The problem with this view is its over-simplification. Creationists define microevolution as change within baraminic kinds and macroevolution as dramatic changes that transform one kind of animal into another. But a quick google search reveals that conventional biology does not share these definitions. To most scientists, macroevolution refers to evolution at or above the species level, while microevolution refers to variations and changes within a species. Do you see the problem? If we assume that created kinds are higher than the species level (they are), then creationists need to incorporate macroevolutionary changes into our creation biology models (i.e. baraminology), because without them, our created kinds are stuck where they are.
Without rapid evolution producing new species, genera, and even families, creation biology is trapped; stuck in a cage of its own making. Creationism requires the origin of species just as much as Darwin did, and most scientists would consider “new species” to be “macroevolution.” Of course, creationists still limit these changes to within created kinds. These terms simply refer to how much a species changes, not how related a species is to all other forms of life.
This Isn’t That Strange
So, the defining question of evolution is not whether “macro” and “micro” evolution occur. It is actually “how fast can ‘macro’ and ‘micro’ changes happen?” This is a subtle adjustment, but not a radical change in belief for creationists. Instead, framing the debate this way actually fits better with a host of other creation arguments. For example, creationists never argue that sedimentation and erosion happen in smaller, “micro” degrees. Instead, we argue that they happen faster than the conventional theory predicts. Why? Because we see so much evidence of vast amounts of sedimentation and erosion in the geological record. We conclude from this data that lots of sedimentation and erosion happened over a geologically short period of time. We also see a staggering number of living and extinct species. Instead of denying macroevolution, why wouldn’t we assume dramatic evolutionary changes can happen quickly, as we have done to geological phenomena? Creationists have traditionally rejected the idea that dramatic morphological changes can occur in nature. But expanding creation biology to include such changes (within kinds) actually fits the mold of current creation arguments.
The Speed of Speciation
The answer to the question above is that macroevolution takes a long time. To argue that speciation within kinds can explain the vast biodiversity of life today in just a few thousand years is, to conventional experts, absurd. Darwin’s proposed mechanism of change, natural selection, takes thousands or else millions of years to split one species into two. The burden of proof is felt on the backs of creationists who must explain what mechanisms can produce such high numbers of species from a few created kinds on Noah’s ark just 4,500 years ago. Such rapid evolution would require observable evolutionary changes occurring on timescales of years to decades! This is a daunting task. As creationist Stephen Lloyd once put it, creationists must “out-evolution the evolutionists.”
It turns out, the creationists were ahead of the curve. Within the last 30 years, evolutionary biologists have suggested something called “contemporary evolution,” which refers to the evolution of traits in contemporary time scales.1,2 What is a contemporary time scale? According to the evolutionists: years to decades! This new theory originated from two scientific fields: evolutionary biology and ecology/conservation. These two areas of study were segregated for many years, assumed to be unrelated because of the vast difference in timescale. But with the advent of contemporary evolution, they have been united for the first time.
We creationists can now use the research of contemporary evolution to build a comprehensive case for dramatic rapid speciation. If speciation can occur on contemporary timescales, the origin of species suddenly appears to fit into a Biblical timeline. So, without further ado, here is the case for rapid speciation.
Premise One: Adaptive Radiation

Before we even touch on observable evidence of rapid speciation, we should start with adaptive radiation. This concept permeated evolutionary biology literature before scientists even began studying contemporary evolution. Adaptive radiation refers to when one species quickly diversifies into many forms in response to a dramatic event.3 For example, when the meteorite allegedly wiped out the dinosaurs 66 million years ago, evolutionists believe the few mammals existing at that time rapidly diversified over the globe, taking over the habitats left by the dinosaurs. This rapid diversification was termed “adaptive radiation.” This should pique the interest of creationists, since this sounds suspiciously like what occurred after the flood of Noah.
Interestingly, conventional literature has largely agreed about the causes of adaptive radiations. This model dates back to George Gaylord Simpson’s book The Major Features of Evolution (1953). His work on the causes of adaptive radiations is foundational to modern speciation theories. He proposed that four factors could cause rapid radiations: 1. the colonization of a new land mass, 2. the appearance of new resources, 3. a great extinction event, and 4. a novel innovation. These four causes can be summarized as “ecological opportunity.”4 5 In other words, when an opportunity arises for a species to fill an available niche (or “ecological space”), it will rapidly colonize the new land and, if necessary, adapt into many distinct forms.
Adaptive Radiation and the Flood
In the evolutionary timeline, adaptive events are few and far between. The appearance of a new landmass might happen once in a few million years, and then an extinction event may occur a few million years later. The theory is designed to explain some sudden bursts in the fossil record, such as when mammals took over the world immediately after the extinction of the dinosaurs. But what does this model mean for creationists? How does this help us get rapid speciation in just a few thousand years?
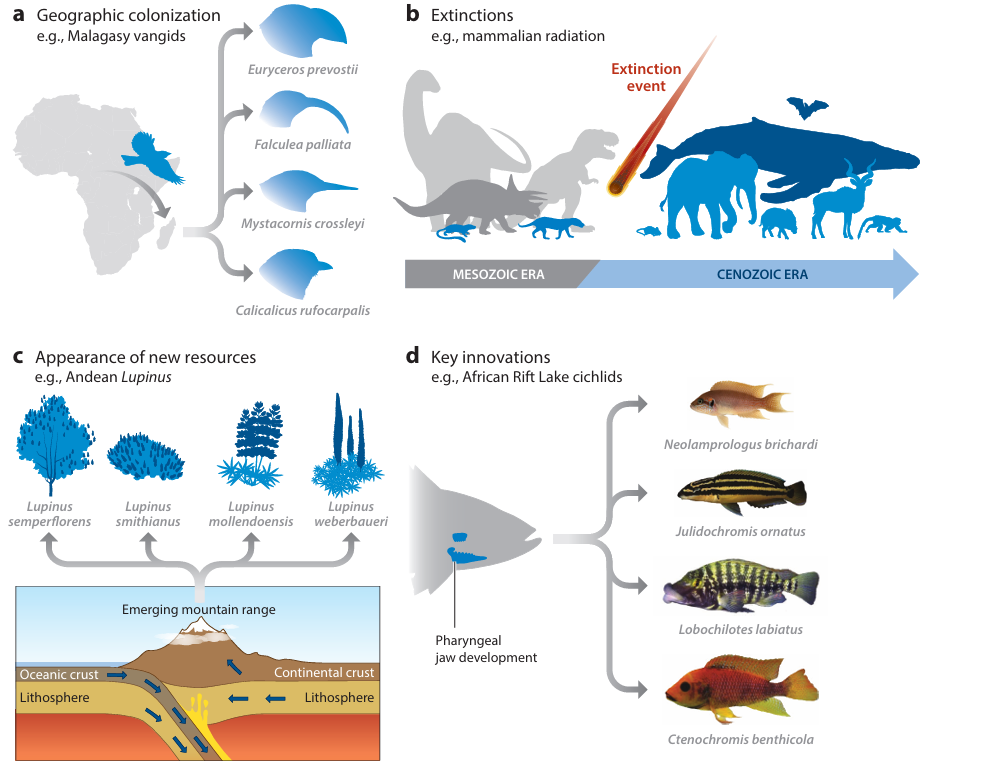
Remember, adaptive radiation almost perfectly describes the type of process that must occur between the Flood and today. So, if there are certain “triggers” that set off an adaptive radiation, creationists would expect to find those triggers right after the flood; ideally to an extreme degree.
Now, take a second to imagine the world right after the Flood. Think of Noah releasing the animals to spread across the globe once again. Now remember GG Simpson’s examples of “ecological opportunity.” Are there new landmasses to colonize? New resources to utilize? Extinction events making room for other species to move in? Each of these phenomena are part of the post-flood world to an astronomical degree! For at the moment the animals began to spread, the entire world was a new landmass, every plant, crevice, and landform is a new resource, and every extinction event (of which there are many) is a catastrophic shift in ecologic opportunity. Imagine the radiations that would happen in a world like that!*
Premise Two: Contemporary Evolution
Recall that the term “contemporary evolution” is a friend of creationism, not a foe. This concept refers to the evolution of biological traits in years to decades.
The greatest critique of creationism’s rapid speciation theory is that there is no well-understood mechanism to explain it. Darwin’s mechanisms are too slow to explain how so many species could come about in just a few thousand years. While this critique is substantial, it is not the death blow to rapid speciation. Observational research has shown rapid speciation happening today! While we might need a mechanism to develop a good model for rapid speciation, we do not necessarily need a mechanism to show that rapid speciation occurs. In fact, when faced with the evidence of rapid evolution today, many evolutionists have proposed new, non-Darwinian mechanisms for it!
These scenarios of contemporary evolution help demonstrate that ecological opportunity drives rapid speciation. Invasive species colonizing a new area often change behavior, morphology, and dispersal ability to fit their new environments. Species colonizing islands also demonstrate rapid diversification due to how many unique resources are available to them. When encountering a new resource, a species will change the rate at which they reproduce, the amount of land they cover, and the time it takes to mate and give birth. By studying examples of invasive species, island colonizations, and migrations, we can start to understand how speciation can happen quickly.
By combining current observational data with Simpson’s “triggers” of adaptive radiation, we can make a strong case that speciation can happen much faster than conventionally thought. Further, we can make this case without knowing for sure what the mechanism is.
An Example of Contemporary Evolution
Observational evidence of contemporary evolution exists in a variety of animals. We’ll give special attention to the vertebrates (backboned animals) that had to spread across the world after the flood.
Our first example takes place in Europe, where a species of migratory bird is rapidly diverging into two species. Due to human activity in the area, the European blackcap has developed two migratory routes. One pathway is the shorter Northwestern route, and the other is longer and Southwestern. This split caused a divide in the species, forming two reproductively separated populations. This division is so strong that when the populations hybridized, their offspring migrated somewhere between the two destinations!6 In addition, the two populations grew morphologically different, with the Northwestern blackcaps having more rounded wings and shorter beaks than the Southwestern blackcaps who have to travel farther.

The study demonstrated that this divide occurred in under 30 generations, making this a clear example of rapid evolution. The paper describing this process suggests that new migratory pathways that result in reproductive isolation can drive rapid speciation. After the flood, the migratory possibilities were endless, as bird kinds discovered migratory pathways in a brand new ecological landscape. This research could demonstrate one way animals speciated after the flood: through reproductive isolation caused by brand new migration routes.
Selective Breeding in Lake Victoria
Another popular study involves cichlid fish in lake Victoria. This example also relies on reproductive isolation (as in, two groups of the same species that do not interbreed). But while the blackcaps required geographical isolation, the same is not the case for the cichlid fish. Instead, these interesting fish divided themselves by color. Female cichlids that had the gene allowing them to see the color red mated only with red fish, and fishes that were missing the gene chose blue mates. Obviously, this will cause a dramatic divergence in the fish population, similar to that of the migratory blackcaps. This genetic isolation is causing contemporary speciation in lake Victoria.7 How fast is this speciation? The research is not clear, but it is happening fast enough to observe and to be relevant to conservation planning.
The implications of this research are potentially wide-ranging. It suggests that wherever there is significant isolation, whether separated by space or a different factor, there is potential for contemporary evolution and thus rapid speciation. A new world has opened up in ecology and evolutionary biology, as biologists begin looking for cases of contemporary evolution.
Darwin’s Tortoises Revisited
In the Galapagos Islands, another study found that tortoises diversified rapidly to adapt to new island environments in a similar fashion as the famous Darwin’s finches. According to research this diversification process “proceeds very rapidly.”8 Though the researchers place these tortoises on the islands hundreds of thousands of years ago, these numbers were based on conventional dating of the volcanic islands in question. The data and interpretation of the research is still consistent with a creation model. The study revealed that rapid evolution occurs both genetically (DNA changes) and morphologically (visible traits change). The interpretation of this data as explained by the researchers is that islands function as modern day examples of ecological opportunity. In other words, this tortoise diversification reflects the power of resources and opportunity to drive forward speciation, building the case for rapid post-flood speciation.
What about Creation Models?
It is worth mentioning that evolutionary models alone cannot explain the rapid speciation problem. The speciation question still requires young-earth scientists to discover what mechanism can cause drastic macroevolutionary change in only a few thousand years. Evolutionists are unlikely to answer this query because they are operating under the assumptions of deep time. They are not trying to fit speciation into a young-earth timeframe. The research presented here demonstrates that processes like adaptive radiation and diversification can help explain the biodiversity seen on earth today. But solving the mechanism for significant morphological change is something creationists will have to do on their own.
To explain this, creationists tend to focus more on the organism rather than the selective pressures placed on populations. In other words, creationists look at an animal as a complex, engineered being, not as an object shaped by external forces. If living things were designed by God, perhaps God engineered them with the genetic ability to change and adapt. This idea has been popular among creationists for a long time, and several biological concepts reflect this idea.
Organism-focused Theories of Change
One such biological concept is that of “plasticity,” which states that a living thing can change its physical characteristics to better adapt to its surroundings, without even altering its genes. This can happen during development (“developmental plasticity”), or during adulthood in general (“phenotypic plasticity”). This is a popular idea in both secular and creationist circles. Taking this a step further, the Institute for Creation Research has proposed a theory called “Continuous Environmental Tracking,” which holds that a creature is always monitoring its own environmental conditions, and can immediately change its internal systems to adapt. While this theory has picked up steam among ICR supporters, other creationists are remain critical and continue to hold more traditional models.
In the end, the organism and its living conditions are both vital components for understanding adaptation and diversification. The data increasingly suggests that speciation is far more complicated than previously thought, leaving the door for rapid speciation wide open.
Conclusion
The case for contemporary evolution is expansive and yet continues to grow. Scientists are unsure about the mechanisms that can cause it, but examples of contemporary evolution are wide-ranging. Models that explain all the data we find are scarce. Most likely, many mechanisms, known and unknown, are involved in the process. It will likely take lots of research to understand the processes that guide speciation. Adaptive radiation, contemporary evolution, and rapid speciation are all different things, but the more we study each one, the more it seems that one can lead to the next. Rapid speciation seems far more plausible than it did just a few decades ago.
Though we rarely see new species arise so quickly today, widespread examples of speedy adaptations and rapid evolutionary change suggest speciation can happen quickly in certain circumstances. It is also likely that the post-flood world matched these circumstances to an extreme.
The exciting part of this creation model is that much of it already exists. The data has been compiled by evolutionary biologists, ecologists, and even conservationists. It only takes a creationist’s careful eye to make it work. We don’t actually need to out-evolution the evolutionists. We might just have to sit back and let them out-evolution themselves.
Notes
*I did not forget about the “novel innovations” part of ecological opportunity. This is an idea that is unconventional in creation biology, but perhaps it was prematurely rejected. Creationists usually don’t hold to the idea that novel traits can evolve, but maybe new mutations contributed to diversification. Let’s table this for now.
Footnotes
- Hendry, A. P., Nosil, P., & Rieseberg, L. H. (2007). The speed of ecological speciation. Functional ecology, 21(3), 455. ↩︎
- Stockwell, C. A., & Ashley, M. V. (2004). Rapid adaptation and conservation. Conservation Biology, 18(1), 272-273. ↩︎
- Stroud, J. T., & Losos, J. B. (2016). Ecological opportunity and adaptive radiation. Annual Review of Ecology, Evolution, and Systematics, 47(1), 507-532. ↩︎
- Stroud, J. T., & Losos, J. B. (2016). Ecological opportunity and adaptive radiation. Annual Review of Ecology, Evolution, and Systematics, 47(1), 507-532. ↩︎
- Myers, E. A., & Burbrink, F. T. (2012). Ecological opportunity: Trigger of adaptive radiation. Nature Education Knowledge, 3(10), 23. ↩︎
- Rolshausen, G., Segelbacher, G., Hobson, K. A., & Schaefer, H. M. (2009). Contemporary evolution of reproductive isolation and phenotypic divergence in sympatry along a migratory divide. Current Biology, 19(24), 2097-2101. ↩︎
- Seehausen, O., Terai, Y., Magalhaes, I. S., Carleton, K. L., Mrosso, H. D., Miyagi, R., … & Okada, N. (2008). Speciation through sensory drive in cichlid fish. Nature, 455(7213), 620-626. ↩︎
- Beheregaray, L. B., Gibbs, J. P., Havill, N., Fritts, T. H., Powell, J. R., & Caccone, A. (2004). Giant tortoises are not so slow: rapid diversification and biogeographic consensus in the Galápagos. Proceedings of the National Academy of Sciences, 101(17), 6514-6519. ↩︎









This was very interesting! I had heard of the cichlid fish a few months back, and it’s great to know that research on them is ongoing. I hadn’t heard of the other examples though! It is so interesting that the blackcaps have changed to such an extreme. I was expecting you to say they changed feather coloration, but that was actual bone and beak structure change. And seemingly instinctual change to, since the offspring of the two populations will just migrate in between the others. That’s not small stuff! Fantastic article and I can’t wait to hear more on all of this!
Thanks Jake for writing this.
I am working on a model of post-flood diversification. This would be consistent with the idea that the fossil record is evidence for post-flood “adaptive radiations”.
With regards to mechanisms for rapid speciation there are some interesting proposals put forward at a recent conference:
https://www.youtube.com/@nordic-creation-research
Thank you for reading! I will take a look.
Hi Jake, I was actually at the CRS conference at MBU just a few days ago and I had a chance to listen to you present this paper in person. After your presentation, I was the guy that asked you to clarify what you meant when you said that you weren’t a macro-evolutionist but also that you believe creationists need to have some allowable framework for some form of macro-evolution. I left your talk chewing over what you had said and have been doing so over the past two days and I think I landed on articulating the essence of my concern. In your presentation, even though you did not define your terms (e.g. “evolution,” “macro-evolution,” nor “evolution-evolution”) all the examples you gave in the paper/presentation were biblically orthodox since they basically reflect a change between Genesis’ kinds. That said, my concern is that since you did not confine the meaning of your terms through definition, I have no idea how to confine my application of your talk into the biological realm. For example, I am clear that you believe in fish-to-fish progression, but am also I to conclude that you also believe in some form of canine to cetacea-like (whale-like) evolution?
Thanks so much,
— Malachi
Hi Malachi, thank you for your questions (both at CRS and here). You are correct that I did not define “evolution,” “macro-evolution,” etc., but doing so was unnecessary in my presentation (hopefully this article cleared up some of that). That is because the topic of my talk was “contemporary evolution,” which I defined as something like “the evolution of new traits in a population within only years or decades.” Then I clarified that contemporary evolution is not the same as molecule-to-man evolution or universal common ancestry. I also explained that creationism NEEDS things like contemporary evolution to explain today’s biodiversity. It was only in the Q&A that “macroevolution” specifically was brought up. In response to a question, I pointed out that the changes that are required to go from two of every kind on the ark to millions of species requires what most scientists consider macroevolution (changes that occur at or above the species level). So, if we use terms as they are typically used in the scientific literature, creationists must accept some form of macroevolutionary change. For example, the differences between a fennec fox and a maned wolf are “macroevolutionary” (they belong to different genera and have vastly different traits, i.e. big ears, furry mane, etc.) but most creationists say they descended from a common ancestor. One dog ancestor producing such different creatures is “evolution” according to most definitions. How powerful are these evolutionary processes? Did land mammals evolve into whales after the flood? I don’t know. But I do believe evolution occurs within created kinds. So, it depends on how broad our created kinds are. For now, I suggest we let the baraminologists figure that out. It is clear to me that creationists already advocate for far stronger and more dramatic evolutionary processes than any evolutionary biologist is willing to consider. But we continue to affirm that God created life within distinct kinds and that universal common ancestry is both false and unbiblical.