

When it comes to extinct animals, dinosaurs capture most of the spotlight. But did you know that in the rock layers below the dinosaurs, we find fossils of another group of animals that were just as wild and varied in size and shape as the “terrible lizards”? These were creatures that were neither reptiles nor mammals, but fell somewhere in the murky middle. What can these creatures tell us about God’s design pattern for animals or the order of burial during the Global Flood?
You are probably already familiar with one of these creatures. You likely recognize Dimetrodon (above left) for its iconic sail formed of skin stretched between long, thin spines sticking up from its back (see image on the left). Its enlarged, conical front teeth and a deep, robust skull made Dimetrodon a fearsome apex predator in its environment.1 Though Dimetrodon is commonly mistaken for a dinosaur, it was actually a type of animal called a synapsid.
The views expressed in this article reflect those of the author, and not necessarily those of New Creation.
What are synapsids?
Synapsids are distinguished from other groups of animals by the number of openings they have in the posterior region of their skulls. Anatomists refer to these openings as temporal fenestrae. A “fenestra” (pl. fenestrae) in anatomy is simply an opening. “Temporal” refers to the temporal region of the skull that the opening “cuts” into. Synapsids are defined as animals that have only one pair of temporal fenestrae (see image below on the right).

All living mammals are classified as synapsids. If you look at an image or model of a human skull, you will see that on either side of the skull, there is a single large opening surrounded by the cheek bones. Your temporalis muscles, which attach to the sides of your skull, pass through these openings to attach to your jawbone. This allows them to close your mouth during chewing. You can think of them as your “nutcracker” muscles. If you put your finger on the side of your head just in front of your ear while opening and closing your jaw, you can actually feel this muscle as it bulges out!
Extinct Synapsids: Trending Towards “Mammalness”
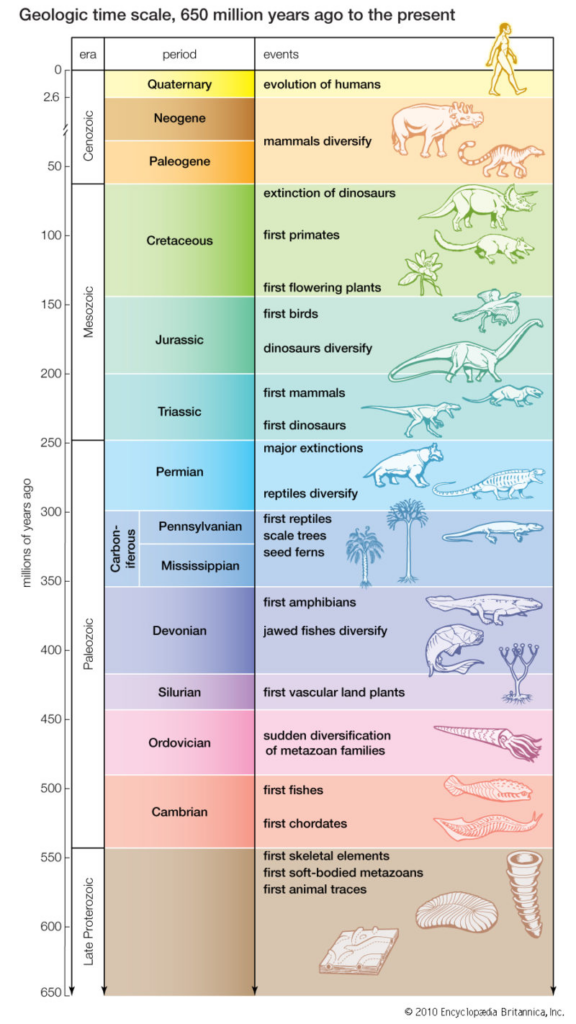
If you look at a skull of Dimetrodon (see the image above), you will see it also has a single pair of temporal fenestrae. This is how we know it is a synapsid. However, if you look at the rest of the skull and skeleton of Dimetrodon, you won’t see much else that is mammal-like. But Dimetrodon is far from the only extinct synapsid we find in the fossil record.
Non-mammalian synapsids (the technical term for all synapsids that are not true mammals) have a wide distribution throughout the fossil record, ranging from the Carboniferous to the Cretaceous systems (see image on the left).2 As we shall see, there is a general trend of increasing “mammalness” as we go up in the fossil record. This trend is punctuated by the sudden appearance of a series of distinct synapsid sub-groups, namely the pelycosaurs, therapsids, and cynodonts.
Pelycosaurs: Sail-Backed Monsters and Tiny Headed Beasts

“Pelycosaurs” are the first synapsids to appear in the fossil record, occurring in rocks from the Upper Carboniferous to Middle Permian systems.3 These animals varied in size as well as diet. Dimetrodon belongs to this subset of synapsids, along with Edaphosaurus, a sail-backed, Dimetrodon-lookalike that was nonetheless a herbivore. Cotylorhynchus was another herbivorous “pelycosaur.” Its massive body could reach twenty feet in length and over a thousand pounds in weight (see image on the right).4,5 Its huge, barrel-shaped ribcage likely housed a large digestive tract for breaking down tough plant matter. Despite its large size, it had a hilariously tiny head.
Therapsids: Terrible Heads and Twin-Tusked Giants
In the Middle Permian, we see the appearance of a second subset of synapsids called the therapsids. Therapsids had features that were more mammal-like than the “pelycosaurs,” including larger temporal fenestrae and a more upright limb posture. Therapsids fit into several distinct groups, such as the dinocephalians and dicynodonts.2

The dinocephalians (“terrible heads”) included massive carnivores like Anteosaurus and herbivores like the gargoyle-headed Estemmenosuchus (see image on the left) and the outlandishly-proportioned Moschops. Moschops had thick, short legs, an enormous, barrel-like chest, and a small, thickened skull ideal for head butting.2

The dicynodonts were herbivorous therapsids that had beaks and often a pair of tusks.2 They are the dominant herbivores in the Permian, though some dicynodonts cross over into Triassic strata. Some were tiny, such as the burrowing, rodent-like Diictodon, while others, like Placerias (see image on the right) and Lisowiscia, could reach the size of bears and elephants, respectively.
Cynodonts: Of Mandibles and Middle Ear Bones

The last group of therapsids we need to discuss are the cynodonts. Cynodonts first show up in the Upper Permian but are more dominant and diverse in the Triassic. Some were small herbivores while others were small-to-medium sized carnivores (e.g., Cynognathus; see image on the left).2
Cynodonts are more mammal-like than all the other non-mammalian synapsids. Their “mammalness” extends to every part of their body, but the most remarkable example can be seen in the relationship between their lower jaws and their ears.
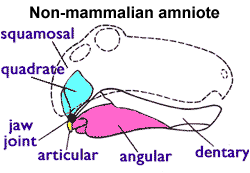
Mammalian jaws are made of a single pair of bones called the dentaries. In contrast, the jaws of reptiles, birds, and non-mammalian synapsids include many other bones. Since these bones are located posterior to the dentaries, they are called postdentary bones (see image on the right).
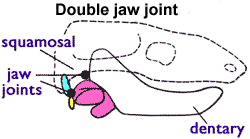
In pelycosaurs and therapsids, the postdentary bones could be fairly large. But in cynodonts, they are very small, located far back on the jaws, and often only loosely connected with the dentary. In the non-mammalian mammaliaforms (the most mammal-like cynodonts) like Morganucodon, this reptile-like jaw joint co-existed with a typical mammalian jaw joint (i.e., the dentaries directly contacted the skull; see image below).
You may remember from biology class that humans have three tiny ear bones commonly called the hammer (malleus), anvil (incus), and stirrup (stapes). This three-piece sound conduction system appears in all mammals. Despite being in our ears, however, the two outer bones (malleus and incus) originate from the same tissue that forms our jawbone during embryonic development. In mice, these bones remain connected to the jawbones (dentaries) by a special type of tissue called Meckel’s cartilage until shortly after birth.6
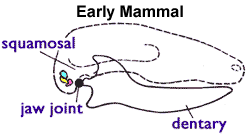
In monotremes (e.g., platypuses), the young hatch at a very early stage in their development, and the middle ear bones remain attached to Meckel’s cartilage until long after birth. The middle ear bones function in supporting the jaw until a connection develops between the dentaries and skull (the typical mammalian jaw joint; see image on the right). This pattern of middle ear bone development exactly mirrors what we see in fossil cynodonts and mammaliaforms.
Synapsids from Evolutionary vs. Creationist Perspectives
Evolutionary Perspective
From an evolutionary perspective, “pelycosaurs,” therapsids, and various non-mammalian cynodonts form a series of successive offshoots from the main branch of the evolutionary tree leading to mammals. Given the clear connection between jaw bones in cynodonts and ear bones in mammals, it is easy to see why they believe this.
Additionally, the order of the fossil record generally fits such a progression. The synapsids furthest in shape from mammals (i.e., “pelycosaurs”) occur in lower strata and those with increasing “mammalness” (i.e., therapsids and cynodonts) occur in successively higher strata until you reach the first true mammals. Such a close alignment between the observed stratigraphic order of fossils and the sequence predicted by evolution is relatively rare in the fossil record.7 Nevertheless, the synapsid fossil record is a clear example of their coincidence. So, are synapsid fossils a knock-out punch against creationism?
Not necessarily. Increasing mammalness is clearly observable in the synapsid fossil record, but what if this pattern actually reflects an underlying blueprint shared between mammals and other synapsids? What if these similarities can be better explained if both shared part of the same master plan originating from the mind of God than by sharing a common ancestor?8
Creationist Perspective
Young-age creationist paleontologist Dr. Matthew McLain has studied non-mammalian synapsids for many years, and he believes this is a large part of the solution.9 He proposes that the real reason that cynodont jaws parallel mammal ear bones is because both have a shared developmental pathway. Mammals are designed to “go all the way” down this developmental pathway to form complete, separated malleus and incus bones, while non-mammaliaform cynodonts were designed to only go partway, with the homologues of these bones remaining connected to the lower jaw.
In his research, Dr. McLain has also found that there are discontinuities separating most of the major therapsid groups from each other. He additionally has found evidence of discontinuity among cynodont groups, including among so-called “primitive” mammaliaforms. In other words, there is no complete, continuous chain linking “pelycosaurs” to therapsids on one end and cynodonts to mammals on the other.10,11,12,13 Instead, most groups appear suddenly in the fossil record already possessing their unique traits. This fits the concept of created kinds and sudden burial during the Flood very well.
But why does the fossil record show such a clear order, with less mammal-like synapsids in lower layers and more mammal-like ones in higher layers? This is a very challenging question to answer, and we can only speculate at this point. It will likely take many years of theorizing and field work before we can give satisfying answers. Nevertheless, given what young-age creationists already think about the structure of the pre-Flood world, Dr. McLain and Dr. Kurt Wise have ventured a possible solution.14
Dr. McLain and Dr. Wise hypothesize that before the Flood, the earth’s environments were highly structured, with different geographic areas housing well-delineated ecological zones. Varying environmental conditions between and within these zones allowed different synapsid groups to thrive in different places. In such a scenario, the degree of “mammalness” in synapsids could have tracked changes in environmental conditions along a gradient across the pre-Flood landscape. If this was the case, it might have been possible for the Flood to have sequentially buried pre-Flood synapsid communities along this gradient, such that the resulting fossil record would show a sequential increase in “mammalness” from bottom to top. This possibility is intriguing, though it is highly speculative. However, support for this idea can be found in the fact that multiple different groups of therapsids show an independent increase in mammalness as you go up the geologic column.9
A lot more work still needs to be done by creationists to solve the synapsid mystery, though some exciting progress is being made! Stay tuned!
Conclusion
Given the current diversity of life on earth, we are used to thinking of mammals as a very distinct type of creature. Compared to living birds, reptiles, and amphibians, mammals are truly unique. But the fossil record shows us an array of strange, now-extinct animals that aren’t quite mammals but aren’t quite anything else either. These “almost-mammals” present a unique set of hard questions for us as young-age creationists to think through. “How do we account for biological similarity from the perspective of design?” “How did the Flood produce the order we observe in the fossil record?” “Why does the order of the fossil record sometimes line up with the order predicted by evolutionary theory?”
While these questions are challenging, they shouldn’t discourage us. Just because we don’t have the full picture now doesn’t mean that we won’t find satisfying answers. We need to continue trusting in the word of our God and King who made all creatures (including synapsids) and do the hard work of rigorous science. With each new discovery, we not only get closer to the answers we seek, but we also get to glimpse the glory and wisdom of our God in what He has made. And that is a prize worth seeking all on its own!
Learn More about “Almost-Mammals”
Footnotes
- Brink, K. S., & Reisz, R. R. (2014). Hidden dental diversity in the oldest terrestrial apex predator Dimetrodon. Nature Communications, 5(1), 3269. https://doi.org/10.1038/ncomms4269
↩︎ - Benton, M. J. (2015). Vertebrate Palaeontology (Fourth ed.). Hoboken, New Jersey: Wiley-Blackwell.
↩︎ - Groups placed in quotation marks are likely not true, organic groupings from an evolutionary perspective. Evolutionists would say these groups are not monophyletic. A monophyletic group includes a common ancestor and all of its evolutionary descendants. If some of the descendants are left out, it is not monophyletic, but paraphyletic. Paleontologists try to avoid using names of paraphyletic groups, except where convenient for communication. ↩︎
- Gould, S. J., & Littlejohn, J. (1973). Factor Analysis of Caseid Pelycosaurs. Journal of Paleontology, 47(5), 886-891. Retrieved from http://www.jstor.org/stable/1303067 ↩︎
- Reisz, R. R., & Fröbisch, J. (2014). The Oldest Caseid Synapsid from the Late Pennsylvanian of Kansas, and the Evolution of Herbivory in Terrestrial Vertebrates. PLoS ONE, 9(4), e94518. doi:10.1371/journal.pone.0094518 ↩︎
- Anthwal, N., Fenelon, J. C., Johnston, S. D., Renfree, M. B., & Tucker, A. S. (2020). Transient role of the middle ear as a lower jaw support across mammals. eLife, 9, e57860. doi:10.7554/eLife.57860 ↩︎
- McGuire, K., Southerden, S., Beebe, K., Doran, N., McLain, M., Wood, T. C., & Garner, P. A. (2023). Testing the order of the fossil record: preliminary observations on stratigraphic-clade congruence and its implications for models of evolution and creation. Proceedings of the International Conference on Creationism, 9, 478-486. doi:10.15385/jpicc.2023.9.1.26 ↩︎
- Wise, K. P., & McLain, M. A. (2025). Common Development as a Synthetic Theory of Creation Biology. Paper presented at the Origins, Dayton, Tennessee. ↩︎
- McLain, M. A. (in review). Therapsids and the Blueprint of Life: A Creationist Approach to Problems of Biological Similarity and the Fossil Record. ↩︎
- McLain, M. A. (2017). Baraminology of Non-Cynodont Therapsids I: “Basal” Therapsids, Biarmosuchians, and Dinocephalians. Journal of Creation Theology and Science Series B: Life Sciences, 7, 4. ↩︎
- McLain, M. A. (2017). Baraminology of Non-Cynodont Therapsids II: Anomodonts, Gorgonopsians, and Therocephalians. Journal of Creation Theology and Science Series B: Life Sciences, 7, 4-5. ↩︎
- McLain, M. A. (2017). Applying Baraminological Methods to Understand Mammal-like Cynodonts. Journal of Creation Theology and Science Series B: Life Sciences, 7, 7-8.
↩︎ - Talavera, A., & McLain, M. A. (2018). Applying Baraminological Methods to Understand “Basal” Mammaliaforms. Journal of Creation Theology and Science Series B: Life Sciences, 8, 4.
↩︎ - Wise, K. P., & McLain, M. A. (2025). Common Development Model for Antediluvian Biozonation and Paleontological Pattern. Abstract presented at the Origins 2025 Conference, Dayton, Tennessee.
↩︎










Is there fossil evidence that cynodonts had whiskers, as in the picture?
Not directly no. However, cynodontsare a group of synapsids that are considered close to mammals. The evidence for whiskers mainly comes from tiny holes in the skull called foramina.
The most complete skulls of cynodont we have shown small openings in the snout/jaw region which are associated with nerves and blood vessels.
In modern mammals, enlarged facial foramina are strongly associated with:
Fossil evidence suggests that some cynodonts may have possessed whiskers, based primarily on facial foramina associated with sensory nerves in the snout. However, no direct fossilized whiskers have been discovered, so the conclusion remains an anatomical inference rather than direct evidence.
Great article and they are one of the best species we have for a holobaramin based on lots of different species I have studied. I really like the new evidence we have where early man drew one in a cave that they saw. Considering these species were only supposed to have lived before dinosaurs even existed, how in the world did mankind ever see one? The only idea they can come up with is that maybe they discovered their fossils, rebuilt them and interpreted the creature accurately then then integrated them into art. However they refute this later by claiming that “Pure imagination may be safely ruled out as the San did not paint things that were completely imaginary,” Benoit explained to Newsweek. “Their art was based on real-life elements, mostly animals.” “Furthermore, San oral traditions include stories of “enormous brutes” that once roamed the land and are now extinct.” To read more check out: Rock Art Suggests Indigenous People Were First Paleontologists
Matt here, I wanted to add another additional explanation to the answer of this question; why does the fossil record show such a clear order, with less mammal-like synapsids in lower layers and more mammal-like ones in higher layers?
I would like to add regarding their geological proximity that another reason we find them distributed as such is because Early synapsids (e.g., Dimetrodon) Had a natural sprawling posture,’making them the least mobile and first to die in a flood.
followed by the more diverse therapsids who had a more upright posture with legs under them. Most species were large, ranged from lion to bear size with many the size of a cow. While larger, they were still more mobile and lived in different echo systems.
Last we have Cynodonts (closest to mammals). They are considered the last to evolve of the chain but consider again limbs and size. Most were small, very small. Typical range: shrew, mouse to the largest bring fox-sized. These are the most mobile of all of them, so logically you would expect them to outrun synapsids and theropods.
Hi Joshua,
I too have spent a number of years researching mammalian evolution and written two articles on the subject for the CMI Journal of Creation.
One conclusion was that Morganucodon did not have a double jaw joint [Is Morganucodon a transitional fossil? 2020 Volume 34 Issue 3]. Also claims of other creatures said to have a double jaw joint and incipient dentary-squamosal joint, have been shown to be incorrect by CT scans of key fossils. [A critical review of the fossil evidence for the origin of the mammalian jaw joint and middle ear, 2025 Volume 39 Issue 2].
I also posted a short video about Morganucodon on my YouTube channel. https://youtu.be/RC8b30U1ehc?si=yaqfvyAD5iVlEnUP
I would like to suggest that you reconsider what you wrote about the temporal fenestra. There is NOT a single large opening on the side of the human skull. The cheek bone is a bar attached to the side of the skull. Mammals do not have a temporal fenestra like that found in true synapsids.
The double jaw joint shown in the diagram “Double jaw joint” would not work, the joints have to have a common axis to allow rotation. A construction as shown would not work. The only way to open the jaw would be to dislocate one of the joints.
Regards,
Marc Surtees