
“Living fossils” represent presently existing organisms that are also found virtually unchanged as fossils in the geologic record. Some examples are the ginkgo family, the sequoia family, horsetails, the Lingula brachiopod, the chambered nautilus, the horseshoe crab, the tuatara lizard and, of course, the coelacanth.
The following article has been reblogged with permission from Creation Unfolding. The views expressed reflect those of the author, and not necessarily those of New Creation.
Recently, however, a number of scientists have sought to remove the term “living fossil” from the paleontological vernacular by appealing to the philosophical underpinnings of evolution itself: Since, according to these scientists, all biological systems continually evolve, all organisms removed from ancient ancestors by millions of years cannot possibly be “close” living descendants of those organisms, even if the descendants bare some morphological affinities to the ancestors. As an example, Casane and Laurenti (2013) say, “we emphasise that concepts such as ‘living fossil’, ‘basal lineage’, or ‘primitive extant species’ do not make sense from a tree-thinking perspective.” These authors then proceed to apply this reasoning to the coelacanth, perhaps the most iconic of all living fossils.
INTRODUCTION
Today, the coelacanth group is made up of just a single genus Latimeria which contains two known species. Comparing this modern genus to the supposedly 400-million-year-old genus Diplocercides, and it doesn’t take long for one to be convinced of their striking similarity, and why the term “living fossil” was aptly applied to the living Latimeria (see figure). And, of course, one can also see why creationists are so interested in this subject. After all, according to standard evolutionary theory, genetic mutations combined with natural selection and genetic drift are constantly at work, changing the genome so that new adaptations and thus new organisms are always evolving.

Of course, evolutionists are not without answers, and several hypotheses have been put forward to explain this remarkable stasis. Some have proposed the work of Stabilizing Selection. This mechanism weeds out the traits of a population that are at one extreme or the other, thus leaving selection to favour those traits representative of the overall average. A popular example is the average weight of human babies (see figure). This average remains constant through time because premature babies, which are small, can struggle to survive while large babies have problems getting through the birth canal. Traits for these two extremes, therefore, tend to get removed from the overall human population, or at least they did just a few hundred years ago before the advent of modern medicine.
This mechanism, however, only works in very stable environments where external pressures are minimized. Yet since the time of Diplocercides, there have supposedly been five colossal mass extinction events with one of them wiping out 95% of all animal taxa alive at that time! On several occasions, huge volcanic eruptions produced thick and extensive sequences of Flood Basalts which covered massive tracts of land and even ocean floor with millions of square kilometres of basalt that simultaneously released trillions of tons of sulphur and carbon dioxide, along with many other harmful gases directly into earth’s oceans. We are also told that the earth has experienced multiple bolide impacts, with the one occurring at the end of the Cretaceous supposedly killing all of the dinosaurs. There is also the collision of Gondwana and Laurentia during the Late Palaeozoic with the subsequent break up of Pangea during the Mid to Late Mesozoic, not to mention the copious swings in climate, oscillating from one extreme to the other over hundreds of millions of years.
Low species diversity has also been hypothesized. Stephen Jay Gould (2002, p. 817), for example, said, “They [living fossils] simply represent the few higher taxa of life’s history that have persisted for a long time at consistently low species number—and have therefore never experienced substantial opportunity for extensive change in modal morphology because species provide the raw material for change at this level, and these groups have never contained many species.” In other words, low numbers of species mean low numbers of mutations which thus means low numbers of useful traits that natural selection can then act upon to build new organisms.
There are, however, a number of problems with this hypothesis as well. The most salient of which is the problem of extinction. If a genus has low species diversity, then that genus as a taxonomic group, is more prone to getting wiped out in an ever changing and competitive environment. Given Gould’s assessment, how in the world could this little subset of fish spend 400 million years dodging multiple mass extinctions, devastating volcanic eruptions, earth shattering bolide impacts, and at least two ice ages? And if that isn’t enough, this little coterie of rather harmless fish somehow avoided being eaten into extinction by an ever-escalating biological arms race that supposedly furnished every other animal in the ocean with more evolved, elaborate, and highly advanced weapons of predatory warfare. As it turns out, and according to a paper published by Bennett et al. (2018), and another published by Toriño et al. (2021), coelacanths may have been more diverse than previously thought anyway.
More recently, researchers concluded that the similarity of form between Devonian coelacanth’s and the modern Latimeria was due to an extremely slow rate of genomic evolution. In other words, coelacanths just didn’t evolve very much. Casane and Laurenti (2013, p. 332), however, point out the difficulties associated with such a hypothesis:
“Genomes change continuously under the combined effects of various mutational processes, that produce new variants, and genetic drift and selection, that eliminates or fixes them in populations. In other terms, the only possibility for genomes to replicate without change implies at least one of the two following conditions: (i) new variants do not appear (i.e. no mutations), and (ii) new variants are systematically eliminated by selection (i.e. no genetic drift and very powerful selection against new variants).”
In other words, how can a genome remain virtually unaffected by mutations for a period of over 400 million years?
Notice that in each one of these solutions, the scientists are appealing to mechanisms that somehow slowed down evolution by huge orders of magnitude. It’s important for creationists to detect the problem here. And it’s not with evolution. Using living fossils to somehow prove that evolution doesn’t happen is not a good approach. In fact, I would say it’s a decidedly bad approach. Natural Selection is a scientific surety, and vast amounts of diversity have certainly occurred since the Flood of Noah. Even in the secular realm, more and more scientific research is affirming that species are diversifying at astonishingly rapid rates. Living fossils, therefore, do not challenge the evolutionary assumption that organisms change. If the Devonian Diplocercides went extinct 100,000 years ago, we wouldn’t be having this conversation. That’s because nothing would be unusual. Secular scientists, however, believe Diplocercides went extinct 400 million years ago. And, for obvious reasons, that presents a problem. And a huge one at that. There is simply no known mechanism that can adequately explain 400 million years of evolutionary stasis in very complex organisms.
The issue then for the evolutionist, is time, not the absence of evolutionary change. The existence of not just one, but of over 600 living genera (Whitmore 2019) that supposedly have 70- to 500-million-year-old fossilized ancestors is, in my opinion, powerful evidence that those ancestors lived not 500 million years ago, but just thousands of years ago.
And this brings us to the fourth explanation put forward by Casane and Laurenti (2013)—coelacanths must have evolved, and so Latimeria, cannot, therefore, be closely related to the Devonian Diplocercides, or in fact to any ancient coelacanth. As such we need to stop calling it a “living fossil.”
In order to make their case Casane and Laurenti compare Latimeria with its closest known relative, Macropoma (middle fish in top figure), which is supposedly 70 million years old. Based on the skeletal data, they argue that Latimeria shows substantial anatomical divergence so as to support their hypothesis that the two genera are in fact not closely related.
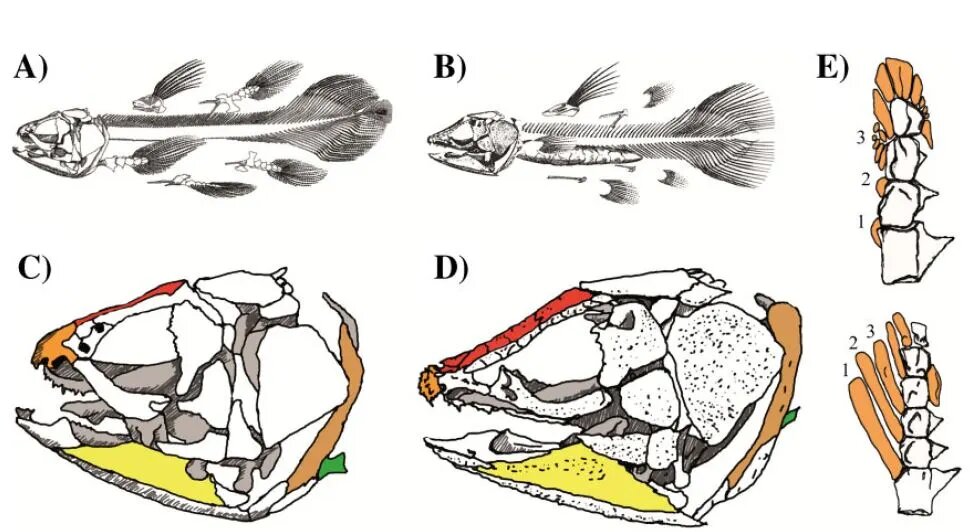
Let me show you why I disagree with their conclusions. Let’s start with the post-cranial anatomy (see figure. Image from Casane and Laurenti 2013. Their figure 2. Open Access). As you can see form their own diagrams Latimeria (left), relative to Macropoma (right), has a longer vertebral mid-section but a shorter post anal region. The hemal arch, which is found at the bottom of each vertebra, has thin spines that run downward. In Macropoma, these spines are longer.
In the skull, Casane and Laurenti point out a number of variances; too many to discuss individually here, but you’ll notice that in nearly all of them, the differences are minor and almost always pertain to aspects of bone thickness, length, slight changes in shape, with slight changes noted in denticle ornamentation on the premaxillary bone:
“In L. chalumnae, the mouth opens upward, the articular bone (yellow) is long and narrow, the parietonasal shield (red) is short, the premaxillary bone (orange) is devoid of denticle ornamentation, the dorsal part of the cleithum (light brown) is spiny, and the scapulocoracoid (green) is located on the ventral side. D: In contrast, in M. Lewesiensis, the mouth opens forward, the angular bone (yellow) is triangular, the parietonasal shield (red) is long, the premaxillary (orange) protrudes and forms a hemispherical snout which is ornamented with prominent denticles, the dorsal part of the cleithrum (light brown) is thick, and the scapulocoracoid, (green) is located more medially.” (Emphasis mine).
In response, it must be pointed out that all of these variations are minor, and none of them would cause us to call into question the very low taxonomic rank of genera shared by both of these taxa. This is very important because the authors are trying to persuade us that Latimeria, when compared to other ancient coelacanths is so different that it cannot be called a living fossil. Yet other palaeontologists disagree. Consider this quote from Modisakeng et al. (2006, p. 479): “Evidence of the minor evolutionary change observed in the coelacanth can be found in its skeleton. The skeleton of the extant coelacanth is almost identical to that of Macropoma” (emphasis mine).
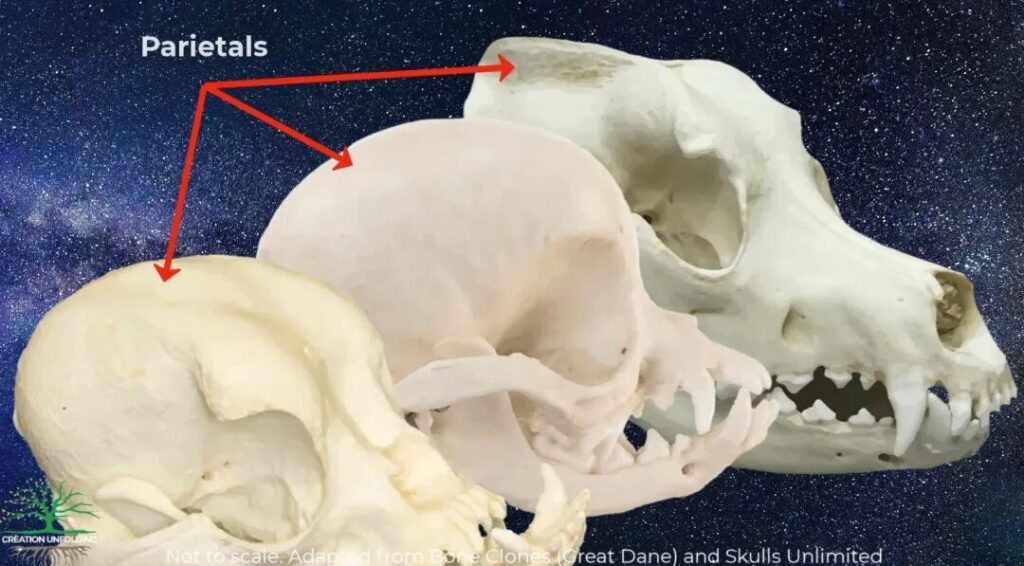
As it turns out, all of the morphological differences that exist between Macropoma and Latimeria can, theoretically exist in the same species, let alone the same genus or family. Consider the variation that we see in different breeds of dogs, all of which consist of a single species. Consider the skulls of an English Bulldog, a Great Dane, and a Chi Wawa (see figure). Notice the incredible plasticity of form that exists just across these three breeds. Every bone in both the neurocranium and the face of each dog is very different. The diversity of form that we see in dog breeds hasn’t escaped the notice of evolutionary researches. Drake and Klingernberg (2010, p. 297), for example, used the skull variation in dog breeds as a proxy for macroevolutionary thinking in higher taxonomic ranks:
“Our analyses show that the variation of cranial shape in dogs is comparable to that across the entire order Carnivora (figs. 3, 4) and that differences in skull shape between extreme dog breeds even exceed the maximal distances we found among the species of Carnivora. This massive disparity among dogs has evolved in a few hundred to several thousand years (Clutton-Brock 1995; Lindblad-Toh et al. 2005), a very brief time span.”
Charles Darwin observed similar degrees of morphological variation among various breeds of pigeons. He said this:
“In the skeletons of the several breeds, the development of the bones of the face in length and breadth and curvature differs enormously. The shape, as well as the breadth and length of the ramus of the lower jaw, varies in a highly remarkable manner. The number of the caudal and sacral vertebrae vary; as does the number of the ribs, together with their relative breadth and the presence of processes. … Altogether at least a score of pigeons might be chosen, which if shown to an ornithologist, and he were told that they were wild birds, would certainly, I think be ranked by him as well-defined species.”
Page 16-17 Darwin’s Origin
Interestingly, this single species of pigeon not only displays, as Darwin puts it, “enormous” differences in the shape and size of individual bones, it also exhibits varying numbers of ribs as well as caudal and sacral vertebrae.
Yes, dogs and Darwin’s pigeons have had their genomes manipulated by human selection pressures, which undoubtedly far exceed the pressures of natural selection. But, in both dogs and pigeons, these dramatic morphological changes occurred in only a few thousand years, and according to Drake and Klingenberg (2010, p. 297), perhaps as low as a few hundred years for dogs. These two coelacanth genera, although not experiencing powerful selection pressures from humans, were supposedly exposed to 70 million years of mutations and natural selection from an ever-changing and, at times, dangerous environment.
In light of these observations, the anatomical comparisons of Macropoma and Latimeria by Casane and Laurenti (2013), actually serve to show that both taxa are surprisingly similar, especially since they have supposedly been evolving for about 70 million years.
Casane and Laurenti (2013) next turn their attention to the pectoral fin of the Devonian coelacanth, Shoshonia (section ‘E’ in figure of skull anatomy above). Shoshonia’s fin is to the bottom and Latimeria’s is at the top. Notice that the first three pre-axial radials are much longer in Shoshonia, than the stubs in Latimeria. In fact, radial three is, in Latimeria, actually broken into a number of smaller pieces and was possibly cartilaginous. The end of the fin in Shoshonia did not preserve and so could possibly have had up to three extra mesomeres, with radials presumably attached to the last one. This is why we don’t see radials at this location in the fossil. Apart, however, from these differences, all of the other skeletal elements in both fins are essentially the same. All of the mesomeres in both taxa are morphologically similar and all are adorned with the same ventral processes. Pre-axial radials four and five, although a little flatter in Latimeria, are still preserved in Shoshonia, as is the post-axial radial along with its semi-lunate shape.
Yes, the two fins are different. But once again we are struck by the similarity rather than the differences, especially when we are told that these fins have been constantly evolving for a period of about 400 million years. The exact taxonomical status of Shoshonia is also questionable as the fossil consists of only a partial fin. Devonian coelacanths were a diverse group and may have been specially created with slightly varying skeletal structures. This is important because this fossil represents the only pectoral skeletal fin for early coelacanths.
Pertinent to this discussion are the findings of other coelacanth scientists. In a paper published in 2012, Zhu et al. (2012, p. 5-6) describe the skull anatomy of yet another Devonian coelacanth. Only in this paper, the scientists are struck by this coelacanth’s affinity with the modern Latimeria:
“As the earliest anatomically modern coelacanth, it [the Devonian coelacanth] shows that the distinctive body plan seen in the living fossil Latimeria has remained basically unchanged from the Early Devonian to modern time. … The finding bodes well for future coelacanth discoveries from the Pragian and earlier strata, and makes the early rapid acquisition of coelacanth characters and the subsequent evolutionary conservatism more impressive than previously thought.”
DISCUSSION
Given all of the above, I propose that Casane and Laurenti’s (2013) claim is unwarranted and that Latimeria retain the appellation, “living fossil.” Interestingly, Bennett et al. (2018, p. 1) have sought to quantify this appellation: “An alternative approach is to use a quantitative metric that matches the most common interpretations of ‘living fossil’ to generate a less subjective listing.” Of course, at the heart of this issue is what’s at stake. Neo-Darwinian philosophy is a powerful belief system and will brook no rivals (see Gould 2002). This mindset can be detected in Bennett et al.’s (2018, p. 2) paper. Although the authors are not targeting creationists, their Neo-Darwinian assumptions come through quite clearly:
“It is argued that the term living fossil is a form of ‘progressivist’ language that can promote a false interpretation of evolution where life is organized into ‘higher’ and ‘lower’ ranks (Rigato and Minelli, 2013). Under a progressivist scheme, these ‘lower’ ranked organisms are then able to subvert post-Darwinian evolutionary thinking by remaining unchanged for millions of years – an impossibility even in the hypothetical circumstance of an absence of selective pressure …”
The term “living fossil” is, therefore, used in various ways and is dependent on the worldview of the person or persons using it. As it turns out Bennett et al. (2018) suggest that the label “living fossil” should be applied to Latimeria. But creationists need to keep in mind that the criteria used for determining what is and what is not a living fossil is itself dependent on the assumptions of those formulating the criteria. Bennet et al. (2018), for example, are motivated by a strict adherence to very modern cladistic methodology.
The term “living fossil” has also been a part of the young-age creationist (YAC) vernacular, especially in the last 60 or so years with the growth of YAC apologetics organizations, books, papers, speakers, and in the last 20 years with the explosion of YAC social media platforms. Its use in this community has traditionally been used as powerful evidence refuting evolution. Yet this kind of thinking is dated, and no longer accords with the scientific evidence. Changes in allele frequencies due to the work of natural selection is a scientific fact. Natural selection in and of itself is observable and measurable. Adding other mechanisms such as mutations and genetic drift to the power of natural selection to produce and fix adaptations may also have some scientific validity. As such, creationists should not use the “living fossil” label as an apologetic refuting evolution.
This of course begs the question: “Then what was the point of this discussion?” The careful reader may already know the answer. All of the secular articles cited in this article ardently uphold evolution’s ever-present, and interminable influence on every organ, tissue, cell, and nucleotide. They may mitigate the rate at which evolution is occurring, but they never, never compromise on its ongoing and perennial work. And neither should they since, as stated above, the science seems to affirm many of these convictions, at least at the micro-evolutionary scale. The specter lurking behind the evolutionary model, then, is not the non-evolution of organisms such as the coelacanth, it is the time over which evolution is supposed to have occurred—it’s way too long.
Think about it, according to evolutionary theory, the Cambrian explosion of multiple complex animals supposedly occurred in as little as 5 million years (Gould 2002). All of the dinosaurs evolved and were exterminated in less than one half of the time that coelacanths have supposedly existed. The whale supposedly evolved from a cow-like mammal in a span of just 8 million years (Gould 2002). Yet the mechanisms of evolution working on the Cambrian organisms, the dinosaurs, and coelacanths are the same. How is it that this complex Devonian vertebrate has remained virtually unchanged in 400 million years? I want you to really think about that. We are not talking about 1000 years. (And as we’ve already seen, lots of changes can occur in just that time). We’re not talking about 100,000 years, or a million years. We are talking about 400 million years. Yet, as this discussion has clearly demonstrated; these two genera (Diplocercides and Latimaria) could just as easily be very close contemporary cousins.
CONCLUSION
In conclusion, all living fossils, but especially those that have ancestors dating back into the Paleozoic, powerfully refute the idea of Deep Time. The most parsimonious and reasonable interpretation of these data suggest that Devonian coelacanths such as Diplocercides lived only thousands of years ago instead of the near half-billion-year timeframe proposed by the secular scientific community.
Learn More about Coelocanths
Footnotes
Bennett, D.J., Sutton, M.D., and Turvey, S.T. 2018. “Quantifying the living fossil concept.” Palaeontologia Electronica, Vol. 21 (1). Doi: 10.26879/750
Casane, D., Laurenti, P. 2013. “Why coelacanths are not ‘living fossils’: a review of molecular and morphological data. Bioessays.” Vol. 35 (4):332-8. doi: 10.1002/bies.201200145. Epub 2013 Feb 4. PMID: 23382020.
Drake, A.G., and Klingenberg, C.P. 2010. “Large-scale diversification of skull shape in domestic dogs: disparity and modularity.” The American Naturalist. Vol. 175(3):289-301. doi: 10.1086/650372. PMID: 20095825.
Gould, S.J. 2002. The Structure of Evolutionary Theory. Cambridge, MA: Belknap Press.
Modisakeng, K.W., Amemiya, Chris., Dorrington, Rosemary., Blatch, Gregory. 2006. “Molecular biology studies on the coelacanth: A review.” South African Journal of Science. Vol. 102.
Toriño, Pablo., Soto, Matías., and Perea, Daniel. 2021. “A comprehensive phylogenetic analysis of coelacanth fishes (Sarcopterygii, Actinistia) with comments on the composition of the Mawsoniidae and Latimeriidae: evaluating old and new methodological challenges and constraints.” Historical Biology: An International Journal of Paleobiology. Vol. 33 (12).
Whitmore, J. 2019. What about Living Fossils? Accessed 01/10/23.
Zhu, M., Yu, X., and Lu, J. (2012). “Earliest known coelacanth skull extends the range of anatomically modern coelacanths to the Early Devonian.” Nature Communications. Vol. 3(772).









